 alexxlab
alexxlab
|
Лидеры продаж ВИКШаблон Красовского УШК-1 Эталоны чувствительности канавочные Магнитный прижим П-образный Фотоальбом дефектов основного металла Альбом радиографических снимков
|

 Микрометры типа МКЦ снабжены цифровым индикатором, повышающим точность и удобство пользования.
Микрометры типа МКЦ снабжены цифровым индикатором, повышающим точность и удобство пользования.
Микрометр. Виды и устройство. Работа и применение. Особенности
Микрометр – это точный измерительный инструмент, предназначенный для работы с деталями мелких размеров. Он обладает высокой точностью, поэтому с его помощью можно получить линейные параметры измеряемого объекта с допуском от 2 мкм. Благодаря столь малой погрешности инструмент и получил свое название. Он намного более точный, чем штангенциркуль, а тем более чем обычная линейка.
Он намного более точный, чем штангенциркуль, а тем более чем обычная линейка.
Существует несколько популярных конструкции микрометров, которые являются усовершенствованной базовой моделью этого инструмента подогнанной под определенные узкие цели.
В простом исполнении микрометр состоит из следующих элементов:
В основе конструкции лежит металлическая скоба, параметры которой ограничивают возможность изменения. На одном ее конце имеется металлическая пятка, а на втором прикрепляется механизм в виде винта. Он отрегулирован таким способом, что расстояние между его кончиком и пяткой скобы отображается на цифровой шкале инструмента. Вкрутив винт до момента прижатия измеряемой заготовки, можно получить точное отображение ее ширины. После этого остается только посмотреть на шкалу. Данный прибор является контактным. Он не применяется для измерения мягких материалов, которые при прикасании начинают сжиматься.
Чтобы полученный результат не сбивался, пока не будет записан, на микрометре предусматривается фиксатор. При его нажатии исключается вероятность случайного выкручивания винтов и сдвига указателя на цифровой шкале даже на несколько долей миллиметра.
При его нажатии исключается вероятность случайного выкручивания винтов и сдвига указателя на цифровой шкале даже на несколько долей миллиметра.
Данное оборудование является довольно распространенным в различных отраслях. Его профессионально используют:
- Токари.
- Литейщики.
- Фрезеровщики.
- Лабораторные сотрудники.
- Моделисты.
- Ювелиры.
Это оборудование позволяет получить точные линейные данные, но оно не столь универсально, как тот же самый штангенциркуль. Для выполнения определенных задач данный инструмент является незаменимым, поскольку именно он позволяет добиться практически лабораторной точности, что не сможет ни один другой ручной прибор измерения.
Виды микрометровСфера использования данного оборудования довольно обширна, поэтому его конструкция была адаптирована под определенные цели. Это позволяет обеспечить максимально удобные и точные измерения. Существуют более 20 конструктивно отличающихся между собой микрометров, из которых многие являются очень редкими и практически не применяются в быту.
Существуют более 20 конструктивно отличающихся между собой микрометров, из которых многие являются очень редкими и практически не применяются в быту.
Среди популярных микрометров можно отметить:
- Гладкий.
- Листовой.
- Для горячего металлопроката.
- Для глубокого измерения.
- Трубный.
- Проволочный.
- С малыми губками.
- Универсальный.
- Канавочный.
- Цифровой.
Самый распространенный в использовании. Он применяется для снятия наружных показателей деталей и заготовок. Именно такой инструмент чаще всего можно встретить в продаже. Подобные модели можно использовать практически в любых целях, кроме тех случаев, когда нужно измерить внутренние показатели заготовок, поскольку для такого устройство не предназначено.
Листовые микрометрыИмеют на пятке и на самом винте круглые тарелки, что увеличивает площадь контакта с измеряемой заготовкой. Это позволяет провести ее предварительную деформацию, чтобы выровнять и измерять точную толщину. Таким инструментом обычно измеряют параметры листового проката, металлических лент и кованых в кузнице заготовок.
Это позволяет провести ее предварительную деформацию, чтобы выровнять и измерять точную толщину. Таким инструментом обычно измеряют параметры листового проката, металлических лент и кованых в кузнице заготовок.
Хотя с теоретической точки зрения снять параметры можно и с помощью обычного гладкого микрометра, но на самом деле это не так. Зачастую прокат имеет неровности, поэтому можно установить пятку и винт на вмятину или наоборот на утолщение. Применение широких тарелок позволяет увеличить площадь и избежать контакта с подобными областями, которые могут приводить к получению неточных данных.
Микрометр для горячего металлопрокатаПрименяется для работы с раскаленными заготовками. C его помощью можно быстро и эффективно измерить толщину железных элементов при их производстве, не ожидая пока они остынут. Именно с помощью этого инструмента удается контролировать момент, когда необходимо остановить прокат металла и забрать готовую заготовку нужных параметров.
Имеют очень вытянутую скобу, которая позволяет накинуть инструмент на заготовку и проверить толщину в удаленном от края месте. Это особенно важно если измеряемая деталь является неравномерной по периметру. С помощью таких устройств можно узнать точную толщину детали, в которой проведено несквозное сверление отверстия или зенкование.
Микрометры трубного типаПредназначены исключения для измерения толщины стенок трубок. Они имеют особенную конструкцию, поэтому их невозможно спутать с устройствами других типов. Визуально определить трубные микрометры несложно. Они имеют обрезанную скобу, на конце которой пятка заменяет срезанную скобу. Такая пятка вставляется внутрь трубки, которая измеряется, после чего винт поджимается и можно получить точные данные о диаметре стенки.
Данное оборудование позволяет снимать параметры даже с очень тонких труб, главное чтобы в них могла войти пятка. Именно это и отличает трубные инструменты от гладких типов. С помощью обычного микрометра можно снимать данные только с довольно толстых труб, внутренний диаметр которых позволяет вставлять в них часть скобы вместе с выходящей в сторону пяткой.
Именно это и отличает трубные инструменты от гладких типов. С помощью обычного микрометра можно снимать данные только с довольно толстых труб, внутренний диаметр которых позволяет вставлять в них часть скобы вместе с выходящей в сторону пяткой.
Является одной из самой компактной разновидностью базовой модели. Он не имеет столь ярко выраженной скобы как обычные инструменты. Внешне его можно принять за обычный металлический прут. Подобный инструмент используется для замера диаметра металлической проволоки и прутиков. Он имеет малый диапазон хода, но этого более чем достаточно для тех измерений, для которых он предназначен. Отсутствие объемной скобы позволяет носить инструмент в компактном чемоданчике с ключами и отвертками. Подобные микрометры занимают места не больше, чем плоскогубцы.
Микрометр с малыми губкамиПредназначен для снятия параметров на поверхности металла после осуществления в нем проточки или сверления. Главная особенность таких инструментов заключается в том, что пятка и винт сделаны очень тонкими. Благодаря этому их можно вставлять в тонкие отверстия. По конструктивным особенностям подобные модели ничем не отличаются от обычных, кроме утонченных элементов.
Главная особенность таких инструментов заключается в том, что пятка и винт сделаны очень тонкими. Благодаря этому их можно вставлять в тонкие отверстия. По конструктивным особенностям подобные модели ничем не отличаются от обычных, кроме утонченных элементов.
Имеют съемные наконечники. Именно такие устройства выбирают в том случае, если нужно проводить измерение, различных по свойствам, заготовок и деталей. Съемные наконечники позволяют адаптировать инструмент под требуемые условия работы. Стоит отметить, что на более дешевых микрометрах данного типа наблюдается одна проблема. При недостаточно сильном зажатии наконечника возможен зазор, влияющий на точность. В том случае если очень точные данные не нужны и погрешность в пол миллиметра не имеет особого значения, то и универсальные модели будут вполне удобными. Приборы более дорогого ценового сегмента зачастую выполнены более качественно, и проблема болтающихся наконечников сведена к минимуму благодаря подгонке всех элементов инструмента.
Предназначены для замера габаритов в труднодоступных местах заготовок. Главной особенностью этого инструмента является полное отсутствие скобы. Внешне они напоминают проволочные модели, но оснащаются специальными тарелками, которые выступают в роли губок, захватывающих детали. С помощью данного оборудования можно зажать выступающие части заготовок губками и измерить их диаметр. Подобные приборы требуют аккуратного обращения, поскольку установленные на их конца тарелочки могут деформироваться при сильном ударе, что случается при падении.
Цифровой микрометрЯвляется одним из самых удобных устройств, поскольку он оснащается электронным дисплеем. С помощью такого оборудования можно намного удобнее и быстрее проводить замеры габаритов деталей заготовок. Питание данного прибора осуществляется благодаря установленной батарейке, такой как используется в наручных часах. По точности они ничем не уступают механическим, хотя и не являются такими долговечными. Электронный дисплей можно разбить, если не относиться к инструменту с достаточной осторожностью.
Электронный дисплей можно разбить, если не относиться к инструменту с достаточной осторожностью.
Более дорогие электронные модели имеют множество кнопок настройки, а также большую встроенную память, поэтому они сохраняют получаемые раннее данные и даже показывают время проведения обмеров. Подобные микрометры будут особенно удобны для промышленного применения, когда необходимо проводить множество измерений в сжатый период времени.
Существует еще как минимум десяток различных типов микрометров. Они являются очень узкоспециализированными, и нельзя сказать, что незаменимыми. Операции, которые они выполняют, можно сделать и другими типами микрометров, что может быть не так и удобно, но точность измерения от этого никак не пострадает. Все микрометры выпускаются в соответствии с требованиями ГОСТ. Для большинства моделей данного инструмента предусматривается отдельный государственный стандарт определяющий точность измерения. Микрометр желательно носить в специальном тубусе, чтобы предотвратить набивания пыли на винт, что убережет его от заклинивания.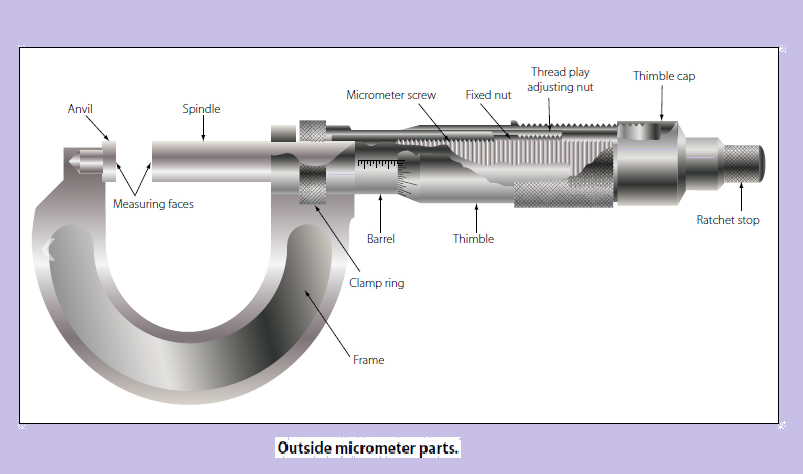
Похожие темы:
назначение и строение конструкции, особенности использования
Функциональное назначение микрометра заключается в измерении относительно малых величин контактным способом. Сферы его применения характеризуются необходимостью получения результатов, отличающихся высокой точностью, например, изготовление инструментов. Инструмент достаточно распространен, однако определенные сложности его применения обусловили актуальность вопроса о том, как пользоваться микрометром.
Конструкция прибора
Современный рынок измерительных приборов предлагает довольно широкий ассортимент микрометров, однако их конструктивное исполнение практически идентично, за исключением моделей электронного типа. Отличия механических приборов заключаются в основном в габаритных размерах измеряемых ими предметов. Стандартный измеритель состоит из следующих компонентов:
- «Скоба». Деталь, представляющая собой основу инструмента, на которой закреплены остальные механизмы прибора.
 Изготавливается из особо прочного металла, устойчивого к деформационным воздействиям, поскольку от жесткости этого элемента напрямую зависит величина погрешности при измерении.
Изготавливается из особо прочного металла, устойчивого к деформационным воздействиям, поскольку от жесткости этого элемента напрямую зависит величина погрешности при измерении. - «Пятка». Элемент, выполняющий функции жесткого упора. Выполняется в двух вариантах: запрессованная в корпус скобы и съемная. Сменная пятка характерна для приборов с диапазоном измерений 500 — 800 миллиметров.
- «Стебель». Составная часть микрометра, выполненная в виде полого цилиндра с размещенной внутри винтовой парой. На лицевой стороне стебля находятся основная, показывающая миллиметры, и дополнительная, показывающая половины миллиметров, шкалы.
- «Барабан». Элемент, шкала которого показывает десятые и сотые доли миллиметра (микрометры), одновременно играет роль указателя для шкалы стебля.
- «Трещотка». Размещена со стороны внешнего торца барабана. Эта деталь не только перемещает микрометрический винт, но и ограничивает величину крутящего момента, прикладываемого человеческой рукой.
 Такая функция обеспечивает правдивость показаний при возникновении упругой деформации элементов винтовой пары и не позволяет повредить механизм прибора.
Такая функция обеспечивает правдивость показаний при возникновении упругой деформации элементов винтовой пары и не позволяет повредить механизм прибора. - «Микрометрический винт». Одно из окончаний элемента имеет гладкую поверхность и выдвигается в измерительную зону, а другое жестко соединено с барабаном.
- «Стопорное устройство». Деталь выполнена в виде винтового зажима, фиксирующего микрометрический винт в момент настройки прибора или снятия показаний.
- «Эталон». Элемент, находящийся вне прибора и предназначенный для его проверки перед проведением измерений.
Класс точности и маркировка
Термин «класс точности» означает максимально допустимую погрешность прибора. Например, максимальная погрешность микрометра «МК25», имеющего первый класс точности, не должна превышать двух микрометров (±0,002миллиметра), тогда как у такого же прибора второго класса — четырех микрометров (±0,004миллиметра).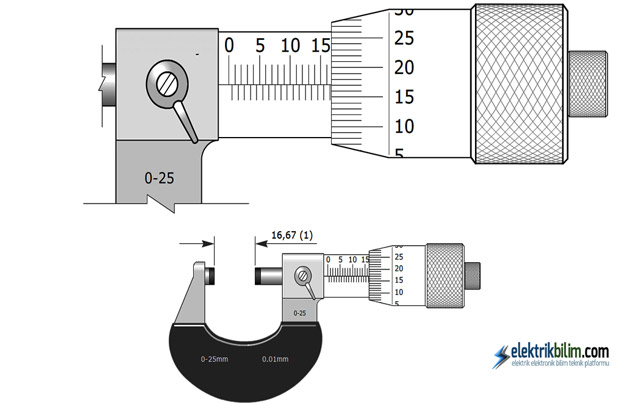
Маркировка измерителя выглядит следующим образом: «Микрометр МК25−1», где число 25 обозначает диапазон возможных измерений (от 0 до 25 миллиметров), а единица — класс точности. Кроме того, к названию добавляется шифр документа, определяющего условные обозначения этих приборов — «ГОСТ 6507−90».
Цифровая индикация
Сегмент измерительных приборов современного рынка инструментов предлагает микрометры, имеющие вместо шкал электронное табло для цифровой индикации измерений. Такие устройства определенно имеют ряд преимуществ в сравнении с их механическими аналогами:
- Цифровое отображение значений значительно упрощает процедуру измерения и минимизирует время считывания показаний.
- Электронные приборы имеют сравнительно малый предел допустимой погрешности и цену деления в один микрометр.
- Цифровые микрометры обеспечивают возможность проведения как абсолютных, так и относительных измерений, что чрезвычайно удобно при проведении технического контроля, выполнении расчетов высокого уровня сложности, разбраковке деталей и тому подобное.
- Способность некоторых приборов «запоминать» пределы допуска.
- Наличие разъема подключения компьютера, позволяющего анализировать статистику измерений с последующим составлением отчетов.
- Возможность использования наряду с метрической системой измерений английскую.
Справедливости ради следует отметить и наличие определенных недостатков, характерных для микрометров с цифровой индикацией измерений. Основной минус — это меньшая в сравнении с механическими приборами надежность, поскольку электронный инструмент более восприимчив к различного рода негативным факторам: ударам, падениям, повышенным температурам и влажности и так далее.
Инструкция по пользованию
Процедура измерения заключается во вращении барабана до момента соприкосновения плоского окончания микрометрического винта и пятки с габаритными окончаниями измеряемого предмета. Поскольку в работе с приборами с цифровой индикацией измерений проблемы возникают редко, рассматривать следует порядок действий на примере микрометра классической конструкции.
Поскольку в работе с приборами с цифровой индикацией измерений проблемы возникают редко, рассматривать следует порядок действий на примере микрометра классической конструкции.
Проверка показаний
Рекомендуется выполнять не только в процессе приобретения прибора, но и постоянно перед выполнением измерений. Процедура проверки начинается с вращения барабана до момента смыкания пятки и плоского окончания микрометрического винта. Прибор работает исправно, если торец барабана останавливается на нулевой отметке шкалы стебля, а продольный штрих указывает на отметку «0» на барабане.
В случае невыполнения одного из условий необходимо произвести регулировку микрометра. Алгоритм выполнения самостоятельной регулировки выглядит следующим образом:
- Посредством стопорного устройства производится фиксация микрометрического винта. Измерительные плоскости при этом находятся в соединенном положении, или между ними зажимается концевая мера.
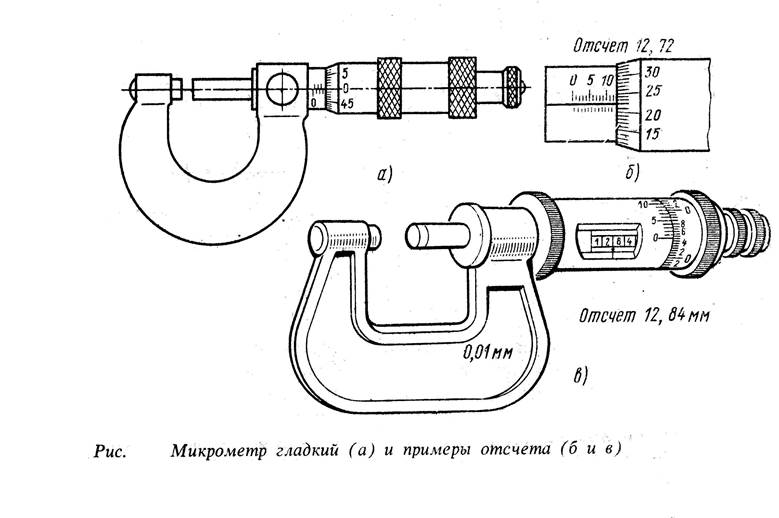
- При помощи специального ключа, входящего в комплект микрометра, выполняется разъединение микрометрического винта и барабана.
- Продольный штрих, нанесенный на стебле, совмещается с нулевой отметкой барабана.
- Прибор собирается в обратном порядке, после чего проверяется повторно.
Фиксация детали
Для проведения измерений деталь должна быть надежно зафиксирована измерительными поверхностями инструмента. Во избежание поломки микрометра и в целях получения максимально точных результатов необходимо придерживаться некоторых простых рекомендаций:
- Плотно прижав измеряемый предмет к пятке, не прилагая усилий, подвести плоскость винта микрометрического к краю предмета.
- Дальнейшее сближение измерительной поверхности винта с габаритом измеряемого предмета производить исключительно посредством трещотки.
- Серия щелчков сигнализирует о соприкосновении измерительных поверхностей с габаритами измеряемого элемента, и показания шкал микрометра соответствуют его размерам.
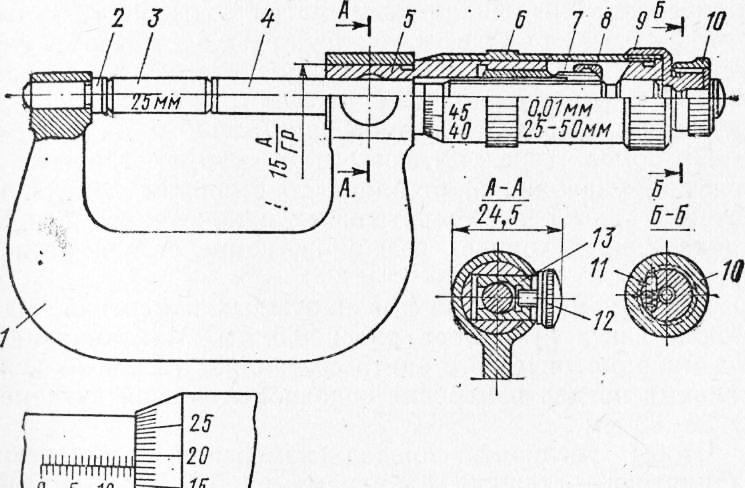
Выполнение этих несложных рекомендаций позволит минимизировать риск повреждения инструмента и существенно снизить степень износа измерительных поверхностей.
Снятие показаний
Снятие показаний начинается с наиболее крупного разряда, постепенно переходя к более мелким. В первую очередь фиксируется показания шкалы, расположенной на стебле. В качестве примера рассматривается модель «МК25−1», цена деления шкалы стебля которого — 0,5 миллиметра. Чрезвычайно важно понимать, что искомый показатель определяется предшествующим открытым делением.
Далее нужно снять показания со шкалы барабана. Здесь цена деления — 0,01 миллиметра. Суммируя полученные показания с двух шкал, получается итоговый результат.
Поверка микрометра
Осуществление поверки микрометра регламентировано методическими указаниями МИ 782−85. Владение методикой поверки чрезвычайно важно как для специалиста, поверяющего инструмент, так и для квалифицированного работника, непосредственно проводящего измерения. Даже в процессе бытовой эксплуатации владение знаниями о поверочных мероприятиях приносит большую пользу. Обнаружение таких отклонений контролируемых параметров, как нарушение параллельности измерительных плоскостей, перекос измерительной плоскости винта и некоторые другие, служат очевидным сигналом о неисправности измерителя.
Даже в процессе бытовой эксплуатации владение знаниями о поверочных мероприятиях приносит большую пользу. Обнаружение таких отклонений контролируемых параметров, как нарушение параллельности измерительных плоскостей, перекос измерительной плоскости винта и некоторые другие, служат очевидным сигналом о неисправности измерителя.
Микрометр: точность измерений без компромиссов
Микрометр – это измерительный инструмент, который сегодня стал незаменимым помощником во многих сферах деятельности человека, вне зависимости от ее направленности. Это легко объяснимо его бескомпромиссной точностью и одновременной простотой в использовании, что делает именно этот измерительный инструмент вариантом выбора специалистов во многих областях промышленности, и не только.
Название этого средства измерения, как и многие другие названия измерительных инструментов, стало производным от единиц измерения, которые легли в основу проведения различных замеров при помощи этого прибора, а именно – микронов. Микрон, как известно, равен одной миллионной доле метра, что обеспечивает максимальную точность любых измерений.
Микрон, как известно, равен одной миллионной доле метра, что обеспечивает максимальную точность любых измерений.
Сфера применения микрометра достаточно обширна. Это средство измерения сегодня применяют:
- в машиностроении;
- в слесарном деле;
- в авто ремонте;
- в токарном деле;
- во многих отраслях строительства и т.д.
Микрометр, цена которого сегодня достаточно демократична, станет незаменимым помощником, если вам необходимо измерить толщину листа железа или другого материала, различных деталей, проводов различного назначения и толщины, проволоки, толщину стенок любых полых элементов деталей и конструкций и т.д.
Актуальность линейных измерений и устройство микрометра
Линейные измерения волновали человека не одну тысячу лет назад и для проведения элементарных замеров, впервые была придумана, знакомая сегодня каждому из нас, линейка. Микрометр, в свою очередь, был изобретен в 1848 году, и в его основу легла микропара «винт-гайка», которую до этого успешно использовали в технологическом процессе производства пушечного механизма. Серийное же производство микрометров, купить которые стало возможным в широкой продаже, началось в США в 1877 году, после того как устройство измерительного прибора было усовершенствовано. Именно такими, какими впервые были созданы микрометры, мы видим из и сегодня.
Серийное же производство микрометров, купить которые стало возможным в широкой продаже, началось в США в 1877 году, после того как устройство измерительного прибора было усовершенствовано. Именно такими, какими впервые были созданы микрометры, мы видим из и сегодня.
Строение микрометра достаточно несложно. Среди конструктивных частей можно отметить следующие:
- D-образная скоба специальной конструкции;
- пятка;
- винт с гайкой;
- шпиндель;
Винт с гайкой в конструкции микрометра занимает место между пяткой и шпинделем и располагаются таким образом, чтобы быть плотно зажатыми между этими деталями. В процессе вращения механизма данного измерительного инструмента, винт фиксируется гайкой, что, собственно, и является основой работы микрометра. Кроме того, микрометр имеет две шкалы, одна из которых расположена на, так называемом, «стебле» микрометра, а другая – представляет собой насечки, расположенный по кругу барабана. В зависимости от класса точности микрометра, первая шкала может иметь цену деления 0,5 или 1 мм, а вторая – иметь 50 или 100 насечек.
В зависимости от класса точности микрометра, первая шкала может иметь цену деления 0,5 или 1 мм, а вторая – иметь 50 или 100 насечек.
Предметом подсчетов, в данном случае, являются полные и неполные обороты винта, которые фиксируются на первой и второй шкале, что и позволяет произвести достаточно точные измерения при помощи микрометра. Точность измерений, которые проведены при помощи микрометра, примерно в 10 раз превосходят аналогичные измерения, проведенные при помощи штангенциркуля и в 100 раз – при помощи обычной линейки.
Такая высокая точность все чаще заставляет специалистов в различных областях купить микрометр вместо штангенциркуля, так как от качества измерений порой зависит не только качество конечной продукции, но и ее работоспособность и последующая пригодность к эксплуатации.
Микрометры механического типа и более современные цифровые модели представлены в широком ассортименте каталога нашего интернет – магазина. Мы предлагаем прямые поставки измерительных инструментов высокого качества от лучших отечественных и зарубежных производителей. Вся продукция сертифицирована и имеет документы соответствующего образца, которые свидетельствуют о высоком качестве и точности любых средств измерения, которые вы можете приобрести в нашем интернет – магазине.
Вся продукция сертифицирована и имеет документы соответствующего образца, которые свидетельствуют о высоком качестве и точности любых средств измерения, которые вы можете приобрести в нашем интернет – магазине.
Микрометр канавочный- 1″ 0,0001″ (0-1″) 146-131 Mitutoyo
Фото может не соответствовать внешнему виду. Производитель оставляет за собой право изменять внешний вид изделия с целью улучшения эксплуатационных характеристик.
Характеристики
| Бренд | MITUTOYO |
| Цена деления, дюйм | 0.0001″ |
| Верхняя граница, дюйм | 1″ |
Описание
Микрометр канавочный- 1″ 0,0001″ (0-1″) 146-131 Mitutoyo – это узкопрофильный инструмент, применяемый в разных сферах промышленности.
33 251
. 40
p
(Без НДС)
40
p
(Без НДС)
39 901 .68 p (С НДС)
В наличии: 10 шт.
В корзину Самовывоз — Москва Доставка по РФ — ПодробнееУченые обнаружили удивительное сходство между строением мозга и Вселенной
https://ria. ru/20201117/mozg-1585005544.html
ru/20201117/mozg-1585005544.html
Ученые обнаружили удивительное сходство между строением мозга и Вселенной
Ученые обнаружили удивительное сходство между строением мозга и Вселенной — РИА Новости, 17.11.2020
Ученые обнаружили удивительное сходство между строением мозга и Вселенной
Итальянские ученые сравнили структуру нейронных сетей человеческого мозга с сетью галактик во Вселенной и обнаружили у них множество общих черт. Результаты… РИА Новости, 17.11.2020
2020-11-17T14:56
2020-11-17T14:56
2020-11-17T14:56
наука
космос — риа наука
физика
нейрофизиология
вселенная
галактики
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdn22.img.ria.ru/images/07e4/0b/11/1585000783_0:32:721:437_1920x0_80_0_0_4447b75a5d5a1e62cfd1717aba1d7581.jpg
МОСКВА, 17 ноя — РИА Новости. Итальянские ученые сравнили структуру нейронных сетей человеческого мозга с сетью галактик во Вселенной и обнаружили у них множество общих черт..jpg) Результаты исследования опубликованы в журнале Frontiers of Physics.Человеческий мозг и Вселенная — две сложнейшие природные системы. Астрофизик Франко Вацца (Franco Vazza) из Болонского университета и Альберто Фелетти (Alberto Feletti), нейрохирург из Университета Вероны предположили, что, несмотря на огромную разницу в масштабе — более 27 порядков, — физические процессы, которые привели к структурированию материи в этих системах, действовали по одним и тем же законам. В итоге сформировались структуры с одинаковыми уровнями сложности и самоорганизации.Человеческий мозг функционирует благодаря наличию обширной нейронной сети, насчитывающей около 69 миллиардов нейронов. Наблюдаемая Вселенная, в свою очередь, состоит минимум из 100 миллиардов галактик. Нейроны и галактики, расположенные в виде длинных нитей и узлов, составляют только около 30 процентов массы систем, а 70 процентов приходятся на компоненты, играющие пассивную роль, — воду в мозге и темную энергию в наблюдаемой Вселенной.Исходя из общих черт, исследователи составили сравнительную модель флуктуации вещества в обеих системах и изучили их структурные, морфологические и сетевые свойства.
Результаты исследования опубликованы в журнале Frontiers of Physics.Человеческий мозг и Вселенная — две сложнейшие природные системы. Астрофизик Франко Вацца (Franco Vazza) из Болонского университета и Альберто Фелетти (Alberto Feletti), нейрохирург из Университета Вероны предположили, что, несмотря на огромную разницу в масштабе — более 27 порядков, — физические процессы, которые привели к структурированию материи в этих системах, действовали по одним и тем же законам. В итоге сформировались структуры с одинаковыми уровнями сложности и самоорганизации.Человеческий мозг функционирует благодаря наличию обширной нейронной сети, насчитывающей около 69 миллиардов нейронов. Наблюдаемая Вселенная, в свою очередь, состоит минимум из 100 миллиардов галактик. Нейроны и галактики, расположенные в виде длинных нитей и узлов, составляют только около 30 процентов массы систем, а 70 процентов приходятся на компоненты, играющие пассивную роль, — воду в мозге и темную энергию в наблюдаемой Вселенной.Исходя из общих черт, исследователи составили сравнительную модель флуктуации вещества в обеих системах и изучили их структурные, морфологические и сетевые свойства. «Мы рассчитали спектральную плотность двух систем. Этот метод часто используется в космологии для изучения пространственного распределения галактик, — приводятся в пресс-релизе Болонского университета слова Франко Вацца. — Наш анализ показал, что распределение колебаний в нейронной сети мозжечка в масштабе от одного микрометра до 0,1 миллиметра следует той же прогрессии, что и распределение материи в космической паутине, но, конечно, в более крупном масштабе — от 5 до 500 миллионов световых лет».Ученые также оценили параметры, характеризующие как нейронную сеть, так и космическую паутину, — среднее количество соединений в каждом узле и тенденцию кластеризации нескольких соединений в центральных узлах внутри сети — и снова выявили неожиданно высокие уровни согласования.»Вероятно, взаимосвязи внутри этих сетей развиваются по схожим физическим принципам, несмотря на поразительную и очевидную разницу между физическими силами, регулирующими распределение галактик и нейронов», — объясняет Альберто Фелетти.
«Мы рассчитали спектральную плотность двух систем. Этот метод часто используется в космологии для изучения пространственного распределения галактик, — приводятся в пресс-релизе Болонского университета слова Франко Вацца. — Наш анализ показал, что распределение колебаний в нейронной сети мозжечка в масштабе от одного микрометра до 0,1 миллиметра следует той же прогрессии, что и распределение материи в космической паутине, но, конечно, в более крупном масштабе — от 5 до 500 миллионов световых лет».Ученые также оценили параметры, характеризующие как нейронную сеть, так и космическую паутину, — среднее количество соединений в каждом узле и тенденцию кластеризации нескольких соединений в центральных узлах внутри сети — и снова выявили неожиданно высокие уровни согласования.»Вероятно, взаимосвязи внутри этих сетей развиваются по схожим физическим принципам, несмотря на поразительную и очевидную разницу между физическими силами, регулирующими распределение галактик и нейронов», — объясняет Альберто Фелетти. «Удивительная степень сходства, которую демонстрирует наш анализ, предполагает, что самоорганизация обеих сложных систем, вероятно, формируется в соответствии с теми же принципам сетевой динамики», — пишут авторы статьи.Ученые надеются, что предложенный ими метод найдет применение как в космологии, так и в нейрохирургии и позволит лучше понять направленную динамику, лежащую в основе временной эволюции как головного мозга, так и Вселенной.
«Удивительная степень сходства, которую демонстрирует наш анализ, предполагает, что самоорганизация обеих сложных систем, вероятно, формируется в соответствии с теми же принципам сетевой динамики», — пишут авторы статьи.Ученые надеются, что предложенный ими метод найдет применение как в космологии, так и в нейрохирургии и позволит лучше понять направленную динамику, лежащую в основе временной эволюции как головного мозга, так и Вселенной.
https://ria.ru/20201111/vselennaya-1584095718.html
https://ria.ru/20201028/mozg-1581892030.html
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2020
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og. xn--p1ai/
xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdn21.img.ria.ru/images/07e4/0b/11/1585000783_40:0:679:479_1920x0_80_0_0_5c8551f1d2bf8865af674c7b53a2e032.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
космос — риа наука, физика, нейрофизиология, вселенная, галактики
МОСКВА, 17 ноя — РИА Новости. Итальянские ученые сравнили структуру нейронных сетей человеческого мозга с сетью галактик во Вселенной и обнаружили у них множество общих черт. Результаты исследования опубликованы в журнале Frontiers of Physics.Человеческий мозг и Вселенная — две сложнейшие природные системы. Астрофизик Франко Вацца (Franco Vazza) из Болонского университета и Альберто Фелетти (Alberto Feletti), нейрохирург из Университета Вероны предположили, что, несмотря на огромную разницу в масштабе — более 27 порядков, — физические процессы, которые привели к структурированию материи в этих системах, действовали по одним и тем же законам. В итоге сформировались структуры с одинаковыми уровнями сложности и самоорганизации.
Астрофизик Франко Вацца (Franco Vazza) из Болонского университета и Альберто Фелетти (Alberto Feletti), нейрохирург из Университета Вероны предположили, что, несмотря на огромную разницу в масштабе — более 27 порядков, — физические процессы, которые привели к структурированию материи в этих системах, действовали по одним и тем же законам. В итоге сформировались структуры с одинаковыми уровнями сложности и самоорганизации.Человеческий мозг функционирует благодаря наличию обширной нейронной сети, насчитывающей около 69 миллиардов нейронов. Наблюдаемая Вселенная, в свою очередь, состоит минимум из 100 миллиардов галактик. Нейроны и галактики, расположенные в виде длинных нитей и узлов, составляют только около 30 процентов массы систем, а 70 процентов приходятся на компоненты, играющие пассивную роль, — воду в мозге и темную энергию в наблюдаемой Вселенной.
Исходя из общих черт, исследователи составили сравнительную модель флуктуации вещества в обеих системах и изучили их структурные, морфологические и сетевые свойства.
«Мы рассчитали спектральную плотность двух систем. Этот метод часто используется в космологии для изучения пространственного распределения галактик, — приводятся в пресс-релизе Болонского университета слова Франко Вацца. — Наш анализ показал, что распределение колебаний в нейронной сети мозжечка в масштабе от одного микрометра до 0,1 миллиметра следует той же прогрессии, что и распределение материи в космической паутине, но, конечно, в более крупном масштабе — от 5 до 500 миллионов световых лет».
Ученые также оценили параметры, характеризующие как нейронную сеть, так и космическую паутину, — среднее количество соединений в каждом узле и тенденцию кластеризации нескольких соединений в центральных узлах внутри сети — и снова выявили неожиданно высокие уровни согласования.
«Вероятно, взаимосвязи внутри этих сетей развиваются по схожим физическим принципам, несмотря на поразительную и очевидную разницу между физическими силами, регулирующими распределение галактик и нейронов», — объясняет Альберто Фелетти.
«Удивительная степень сходства, которую демонстрирует наш анализ, предполагает, что самоорганизация обеих сложных систем, вероятно, формируется в соответствии с теми же принципам сетевой динамики», — пишут авторы статьи.
Ученые надеются, что предложенный ими метод найдет применение как в космологии, так и в нейрохирургии и позволит лучше понять направленную динамику, лежащую в основе временной эволюции как головного мозга, так и Вселенной.
28 октября 2020, 11:36НаукаУченые описали повреждения мозга, связанные с COVID-19Микрометр окулярный — Справочник химика 21
Окулярный и объективный микрометры. Для определения размеров кристаллов необходимы окулярный и объективный микрометры. Окулярный микрометр представляет собою круглую стеклянную пластинку (иногда в металлической оправе), на которой на равных расстояниях нанесены деления (рис. 37). [c.36]
Может быть применен также микроскопический метод для определения дисперсности эмульсии.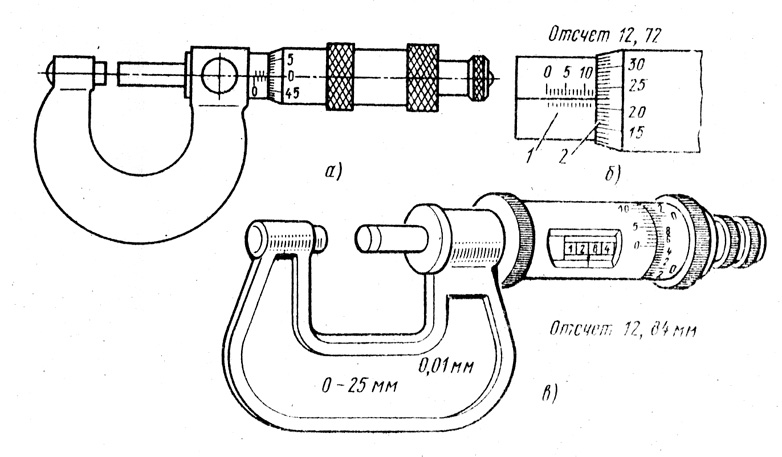 При помощи микроскопа можно определить размеры отдельных частиц, применяя специальные измерительные приспособления, например окулярный микрометр (рис. 12, 13). Однако по этому методу нельзя получить точных результатов, так как практически измерению подвергается лишь незначительная часть имеющихся в эмульсии частиц. Кроме того, при микроскопическом анализе эмульсий нельзя избежать ошибок, получаемых вследствие испарения жидкости в тонком слое, а также деформации частиц покровным стеклом. Поэтому микроскопический дисперсный анализ менее надежен и его можно применять, главным образом, для качественной характеристики эмульсий. [c.28]
При помощи микроскопа можно определить размеры отдельных частиц, применяя специальные измерительные приспособления, например окулярный микрометр (рис. 12, 13). Однако по этому методу нельзя получить точных результатов, так как практически измерению подвергается лишь незначительная часть имеющихся в эмульсии частиц. Кроме того, при микроскопическом анализе эмульсий нельзя избежать ошибок, получаемых вследствие испарения жидкости в тонком слое, а также деформации частиц покровным стеклом. Поэтому микроскопический дисперсный анализ менее надежен и его можно применять, главным образом, для качественной характеристики эмульсий. [c.28]
Крахмал. Готовят два препарата — в растворе йода (раствор Люголя) и в воде от йода крахмальные зерна окрашиваются в синий цвет. В воде определяют их форму, строение, размеры крахмальных зерен измеряют окулярным микрометром. [c.279]
При определении размеров частиц с помощью микроскопа применяют окулярный микрометр, представляющий собой окуляр микроскопа со вставленной в него измерительной щкалой (рис..jpg) 1,а). Цена деления окулярной шкалы зависит от соотношения увеличений окуляра и объектива микроскопа и должна быть определена предвари-, тельно. Для этого на пред- метный столик микроскопа помещают предметный [c.7]
1,а). Цена деления окулярной шкалы зависит от соотношения увеличений окуляра и объектива микроскопа и должна быть определена предвари-, тельно. Для этого на пред- метный столик микроскопа помещают предметный [c.7]
Перед измерением объективный микрометр кладут на столик микроскопа, ставят нужный объектив и окуляр с окулярной линейкой и наводят на резкость, чтобы деления линейки объективного микрометра были четко видны. Далее совмещают в горизонтальном положении обе линейки и высчитывают величину одного деления линейки окулярного микрометра. Например, 50 делений окулярного микрометра линейки соответствуют 30 делениям линейки объективного микрометра, одно деление которой равно 10 мкм. Значит, 50 делений линейки окулярного микрометра равны 300 мкм, а одно ее деление — 6 мкм (300 мкм 50). [c.376]
Прямоугольная рифленая пластинка 3 подвешена с помощью жесткой нити 4 к пружинному динамометру 6. Пластинку полностью погружают в исследуемую дисперсную систему, помещенную в кювету 2 до начала испытания. Кювету С дисперсной системой закрепляют на подъемном столике I. При опускании с постоянной скоростью столика с кюветой пружина растягивается и в системе возникает напряжение сдвига, которое, очевидно, пропорционально растяжению пружины. Последнее может быть измерено с помощью микроскопа 5, снабженного окулярным микрометром, или с помощью микрошкалы. [c.334]
Кювету С дисперсной системой закрепляют на подъемном столике I. При опускании с постоянной скоростью столика с кюветой пружина растягивается и в системе возникает напряжение сдвига, которое, очевидно, пропорционально растяжению пружины. Последнее может быть измерено с помощью микроскопа 5, снабженного окулярным микрометром, или с помощью микрошкалы. [c.334]
Цена деления окулярного микрометра зависит от увеличения данной оптической системы. При замене одного окуляра или объектива другим и при изменении длины тубуса микроскопа (т. е. расстояния между окуляром и объективом) цена деления микрометра меняется. [c.64]
Окулярным микрометром прибора УПОИ-6 определяют длины нарезанных лунок на гильзах цилиндров, затем вычисляют глубины этих лунок по формуле [c.40]
По такому же принципу градуируют окулярную шкалу и в металлографическом микроскопе, только в этом случае используется непрозрачный объект-микрометр. [c.115]
[c.115]
Для осуществления дисперсионного анализа грубых дисперсий широкое применение в практике находит так называемый ситовой анализ, основанный на последовательном просеивании порошка через набор сит с отверстиями определенного, все уменьшающегося размера. Таким образом, удается разделить порошок на ряд фракций с частицами, величина которых лежит в определенных пределах. В случае грубых дисперсий определение величины частиц возможно при помощи микроскопа, снабженного окулярным микрометром. [c.313]
Рис 71 Окулярный микрометр Мея, применяемый для счета н определения размеров частиц [c.228]
Затравочные кристаллы представляли собой плоскогранные монокристаллы размером от 5-10″ до 10 м, полученные спонтанным методом. Расположение кристаллов в реакционном объеме фиксировалось до и после опыта, поскольку их смещение от исходного положения за счет деформации контейнера при сжатии может быть значительным. Величина наросшего и растворенного слоя измерялась с использованием окулярного микрометра МОВ 1-15 с ошибкой 5-10 м.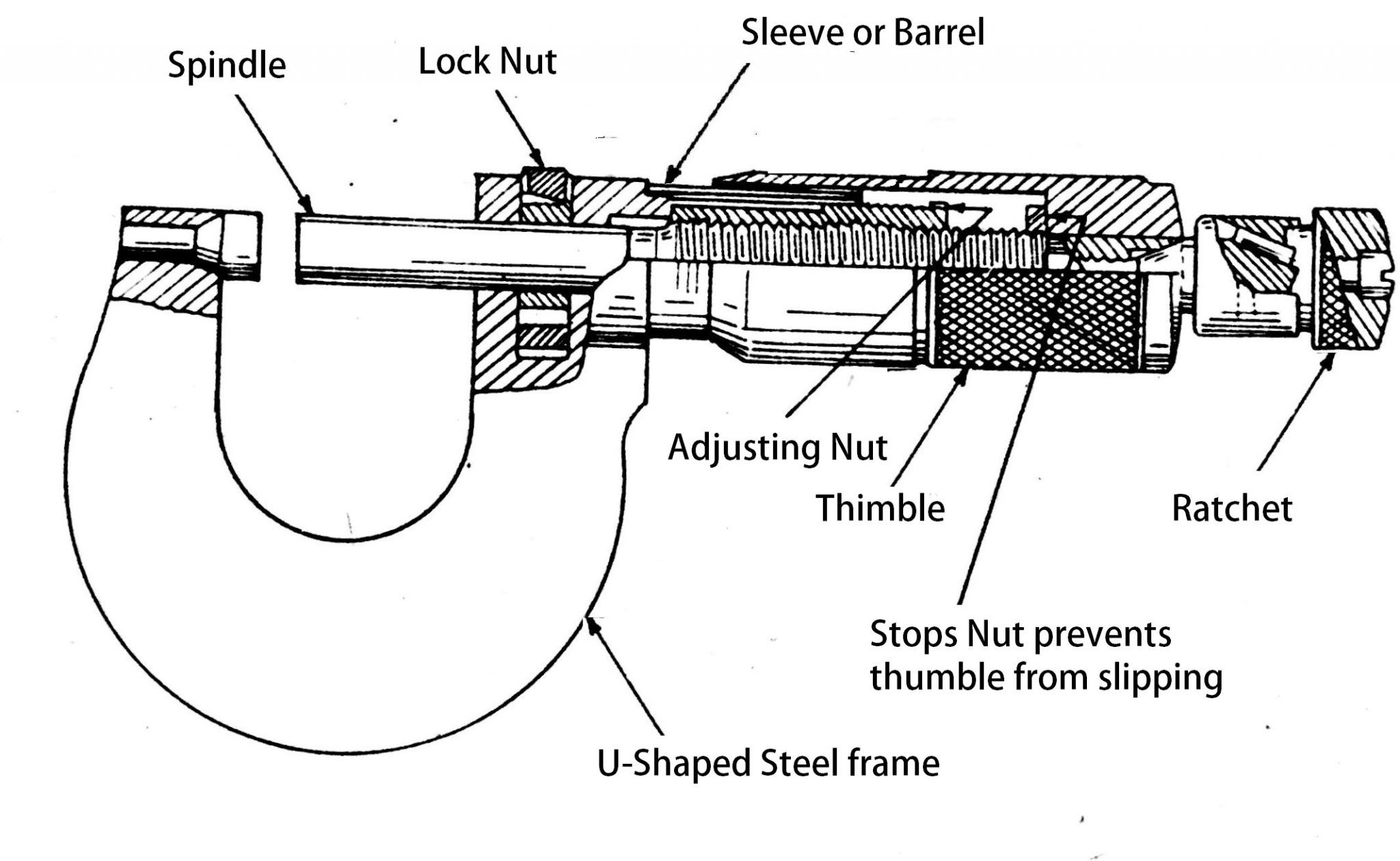 [c.383]
[c.383]
Для этого определяют, какому числу делений окулярной линейки соответствует величина измеряемого объекта, и умножают это число на цену деления окулярного микрометра. [c.16]
Объективный микрометр представляет собой специдльное предметное стекло с размещенной на нем линейкой длиной в 1 мм, имеющей 100 делений, каждое из которых соответствует 10 мкм или 0,01 мм. Окулярный микрометр — это круглое стекло, которое помещают в окуляр микроскопа. На него также нанесена линейка длиной 0,5 или 1 см, разделенная соответственно на 50 или 100 делений. Величина одного деления этой линейки зависит от системы микроскопа, увеличения окуляра, бинокулярной насадки и пр. Поэтому при измерении объектов для каждого микроскопа и для различных увеличений, с которыми приходится работать, нужно определить значение одного деления окулярной линейки в микрометрах (мкм). [c.376]
Измерив объект с помощью линейки окулярного микрометра, умножают число делений на значение этой величины в микрометрах при данном увеличении.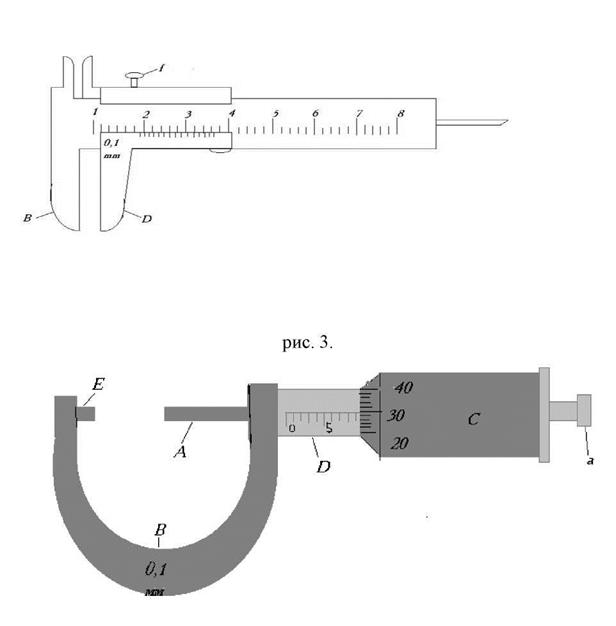 Яйца гельминтов измеряют при большом увеличении микроскопа (объектив 40х). Перед исследо- [c.376]
Яйца гельминтов измеряют при большом увеличении микроскопа (объектив 40х). Перед исследо- [c.376]
Оборудование паразитологической лаборатории почти не отличается от оснащения бактериологической лаборатории и включает, помимо него, окулярный микрометр и нагревательный столик к микроскопу. Препараты кала следует готовить в вытяжном шкафу. Отработанный материал обезвреживают кипячением, стерилизацией или дезинфицирующими растворами. [c.442]
На рис. .26 нанесены в виде четырех точек значения пути разрыва в зависимости от времени, найденные в опытах (см. рис. .25). Светлыми точками помечены результаты опытов определения скорости разрыва с помощью наклеенных алюминиевых полосок. Для пуска искровой аппаратуры использовали пять алюминиевых полосок. С помощью генератора колебаний с собственной частотой 1,7 МГц (следовательно, с точностью измерения времени выше чем 10 с) измеряли время прохождения разрыва между двумя следующими друг за другом полосками.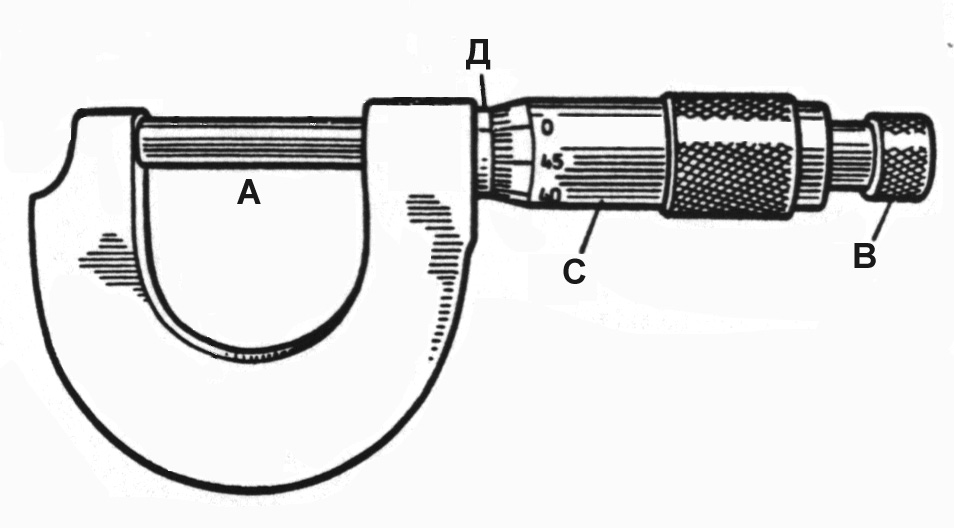 Кроме того, в микроскопе с окулярным микрометром можно было очень точно [c.275]
Кроме того, в микроскопе с окулярным микрометром можно было очень точно [c.275]
Измерительные микроскопы (в том числе портативные накладные) содержат набор измерительных шкал, расположенных в плоскости изображения микрообъектива и позволяющих контролировать линейные размеры деталей, радиусы, углы заточки резцов и т.п. Точность измерения с помощью окулярного винтового микрометра типа МОВ-15 достигает 0,5. .. 1 мкм при увеличениях 10 . .. 20 . Поле зрения микроскопов обычно 1. .. 20 мм. Многие измерительные микроскопы оборудованы устройствами прецизионного перемещения изделий в предметной плоскости микрообъектива с возможностью отсчета координат. Это расширяет диапазон измерений при сохранении высокой точности (диапазон перемещения составляет 50. .. 200 мм, погрешность отсчета — до 1 мкм). Увеличение и соответственно глубина резкости микроскопов выбираются, исходя из особенностей формы изделий. Многие модели современных измерительных микроскопов снабжаются устройствами измерения вертикального перемещения микрообъектива, т.![]() е. обеспечивается трехмерное измерение объектов. [c.491]
е. обеспечивается трехмерное измерение объектов. [c.491]
При этом наблюдают полосы интерференции, искривленные в соответствии с профилем исследуемой поверхности. Кривизну полос измеряют, например, с помощью окулярного микрометра. Она является количественной характеристикой микронеоднородностей поверхности изделия. Измерения можно проводить в белом или монохроматическом свете. [c.498]
Тонкость отсева может быть непосредственно определена микроскопическим анализом и, косвенно — седи-ментациоиным анализом фильтрата. Несмотря на достоинства пер1В0Г0 метода, как прямого способа измерения, он применяется ограниченно, вследствие своей трудоемкости, которая усугубляется при малой концентрации частиц в фильтрате. Для анализа пригоден наиболее распространенный тип учебного, биологического микроскопа с 600-кратным и меньшим увеличением. Капля исследуемой суспензии наносится на предметное стекло и закрывается покровным стеклом. В качестве предметного стекла удобно использовать камеру Горяева или Бюркера, которые применяются в практике медицинских исследований, и обеспечивают толщину рассматриваемого слоя суспензии 0,1 мм. Крестообразный столик СТ-5, в держателях которого закрепляется предметное стекло, и вместе с которыми оно может перемещаться в двух направлениях, позволяет просматривать в проходящем свете последовательно отдельные участки слоя суспензии. В окуляр микроскопа предварительно помещается окулярная сетка — стекло с нанесенной на него сеткой. Цена деления окулярной сетки при выбран-НО.М увеличении микроскопа определяется по объект-микрометру, помещаемому на предметный столик микроскопа. Цена деления на стекле объект-микрометра 0,01 мм. [c.43]
Крестообразный столик СТ-5, в держателях которого закрепляется предметное стекло, и вместе с которыми оно может перемещаться в двух направлениях, позволяет просматривать в проходящем свете последовательно отдельные участки слоя суспензии. В окуляр микроскопа предварительно помещается окулярная сетка — стекло с нанесенной на него сеткой. Цена деления окулярной сетки при выбран-НО.М увеличении микроскопа определяется по объект-микрометру, помещаемому на предметный столик микроскопа. Цена деления на стекле объект-микрометра 0,01 мм. [c.43]
Величину АЛ оценивают нг глаз или измеряют окулярным винтовым микрометром. [c.498]
Для определения размеров некоторых объектов под микроскопом (длина и ширина разновесок, диаметр капилляра и т. д.) необходимо, кроме микроскопа, иметь окулярный и объективный микрометры. [c.64]
Окулярный микрометр представляет собою стеклянный диск (иногда в металлической оправе), на котором на равных расстояниях нанесены деления. [c.64]
Объективный микрометр помещают на предметный столик микроскопа таким образом, чтобы начальный штрих его шкалы совпадал с начальным штрихом шкалы окулярного микрометра, и определяют, какому числу делений объективного микрометра соответствует определенное число делений оку-, лярного микрометра. В предлагаемом на рис. 60 примере всей длине окулярного микрометра при данном увеличении соответствуют 47 делений объективного микрометра, или 470. ш. Значит, одно деление окулярного микрометра при данном увеличении соответствует 4,7 мк. [c.65]
Ошибка измерения не превышает половины цены деления шкалы. Чем больше увеличение, тем меньше цена деления окулярного микрометра и тем меньше ошибка измерения. [c.65]
Параметры /гис/ определяют с помощью окуляр-микрометра 3. В фокальной плоскости окуляра виитового окуляр-микрометра установлены неподвижная окулярная шкала, разделенная на 8 делений, и подвижная шкала с перекрестием и индексом в виде двух параллельных штрихов, расположенных точно над перекрестием. Подвижная шкала приводится в движение вращением барабана микрометрического [c.23]
В верхней части лапки делают прорез 1, затем в верхней и нижней частях просверливают отверстия с винтовой нарезкой, через которые ввинчивают штифт 2, укрепляющий микроскоп 3 в горизонтальном положении. Передвигая лапку по штативу, делают грубую наводку микроскопа. Тонкая наводка достигается поворотом барашка 4. В окуляр микроскопа вставляют окулярный микрометр 5 ДЛЯ измерения отклонения нити ультрамикровесов. Чтобы регистрировать небольшие смещения нити, увеличение микроскопа должно быть 50—80-кратным. [c.69]
Определение толщины прозрачных лаковых покрытий. Для измерения толщины лакокрасочных покрытий на деревянных подложках непригодны магнитные толщиномеры, широко применяемые при измерении толщины покрытий на металлических подложках. МЛТИ совместно с ВПКТИМ разработан метод определения толщины прозрачного лакового покрытия на дереве с помощью двойного микроскопа МИС-11 или микроскопа ППС-2 и микрометра окулярного винтового МОВ-1-15 по ГОСТ 7865—56. [c.110]
На окулярных микрометрах имеется 10 или только 6—8 делений, в свою очередь разделенных на 10 частей. Окулярный микрометр помещается на специальную вставку, имеющуюся в каждом окуляре. Вставка в виде кольца сделана из жести или латуни и может быть 0(пущена на нужную глубину в цилиндр окуляра. Глубина должна быть такой, чтобы при наблюдении через верхнюю. пинзу окуляра были четко видны линии и цифры на микрометре. [c.64]
При опускании с постоянной скоростью столика Е с кюветой пружина Д растягивается, и в системе возникает напряжение сдвига Р. Р астяжение пружины измеряется с помощью микроскопа М, снабженного окулярным микрометром, или по шкале 1Л. [c.234]
Полоску подвешивают к пружинным микровесам 2, изготовленным из молибденовой проволоки толщиной 0,10—0,15 мм. Микровесы помещены в широкую стеклянную пробирку 3 с внутренним диаметром 25 мм. Пробирка шлифом может быть соединена с испарителем 7. Потеря веса образца фиксируется микроскопом 5 с винтовым окулярным микрометром 6 по изменению положения визира 4. Для термостатироаа шя полоску стеклоткани с исследуемым образцом смазки помещают в испаритель 7, через который пропускают сухой воздух или инертный газ (азот, аргон), предварительно нагретый до температуры оиыта в спиральной трубке 10. Скорость воздуха контролируется реометром 1. Температуру в испарителе измеряют термопарой [c.365]
Измерение яиц гельминтов. При идентификации яиц гельминтов в некоторых случаях нужно учитывать их размеры. Так, только по размерам можно отличить яйца фасциолы, гигантской фасци-олы, анкилостомы, некатора, трихостронгилид и пр. (рис. 9.2). Величину яиц гельминтов определяют под микроскопом с помощью объективного и окулярного микрометров. [c.376]
При измерении длины (ширины, диаметра) какого-нибудь объекта под микроскопом его помещают на предметное стекло, совмещают одну из крайних точек объекта с нуловым делением окулярного микрометра и отсчиты.вают, сколько делений Ш1калы занимает измеряемый объект. [c.65]
микрометров | Типы и характеристики измерительных систем | Основы измерения
Микрометр — это инструмент, который измеряет размер цели, закрывая ее. Некоторые модели даже могут выполнять измерения с точностью до 1 мкм. В отличие от ручных штангенциркулей, микрометры соответствуют принципу Аббе, который позволяет им выполнять более точные измерения.
В общем, термин «микрометр» относится к внешним микрометрам. Также существует множество других типов микрометров в зависимости от различных измерительных приложений.Примеры включают внутренние микрометры, микрометры внутреннего диаметра, микрометры трубы и микрометры глубины. Диапазон измерения различается каждые 25 мм, например от 0 до 25 мм и от 25 до 50 мм, в зависимости от размера рамки, поэтому необходимо использовать микрометр, соответствующий цели. В последнее время невероятную популярность приобрели цифровые микрометры.
Аббе гласит: «Чтобы повысить точность измерения, цель измерения и шкала измерительного прибора должны располагаться коллинеарно в направлении измерения.«В микрометрах шкала и положение измерения коллинеарны, поэтому эти инструменты следуют принципу Аббе. Таким образом, можно сказать, что микрометры обладают высокой точностью измерения.
- А
- Наковальня
- В
- Шпиндель
- С
- Зажим
- D
- Гильза
- E
- Рама
- F
- Плита термостойкая
- G
- Масштаб
- H
- Наперсток
- I
- Ограничитель с храповым механизмом
- Поместите цель между наковальней и шпинделем, а затем поверните наперсток, чтобы зафиксировать цель между двумя поверхностями.
- Перед измерением протрите поверхности опоры и шпинделя чистой тканью. Это удаляет грязь и пыль с поверхностей, что позволяет проводить точные измерения.
- Чтобы удерживать микрометр, удерживайте термостойкую пластину на раме большим и указательным пальцами левой руки и зажмите наперсток между большим и указательным пальцами правой руки.
- Возьмитесь за цель между наковальней и шпинделем, поверните ограничитель храповика до проскальзывания, а затем прочтите значение.
- Считайте значение по основной шкале на рукаве и шкале на наперстке. Используйте линию на правом краю рукава, чтобы прочитать значение с шагом 0,5 мм. Затем вы можете использовать шкалу, по которой проходит центральная линия наперстка (шкала), чтобы считать значение в единицах 0,01 мм.
- А
- Шкала наперстка показывает «0,15».
- В
- Шкала наперстка превышает «12».0 мм «
- Используйте измерительный блок или специальный измерительный прибор для калибровки микрометра. Для точных измерений поверхность наковальни всегда должна быть плоской. После выполнения нескольких измерений поверхность может перестать быть плоской из-за износа и накопления грязи. Поэтому периодически используйте деталь, известную как оптическая плоскость, чтобы проверять, является ли поверхность плоской в соответствии с отображаемыми кольцами Ньютона.
- При измерении металлической цели и выполнении калибровки с помощью измерительного блока соблюдайте осторожность в отношении теплового расширения.По возможности не держитесь за металл голыми руками или используйте перчатки, не пропускающие тепло и предназначенные для точной работы.
- Межповерочный интервал микрометров составляет от 3 месяцев до 1 года.
ИНДЕКС
Конструкция винтового датчика микрометра
Конструкция винтового датчика микрометра
Конструкция микрометра с винтовым калибром
На этом изображении вы можете найти структуру микрометрического винта калибра.
Наш ПОСЛЕДНИЙ YouTube-фильм готов к запуску. Просто взгляните, оставьте свой ценный совет, дайте нам знать и подпишитесь на нас! Большое спасибо!
Мы рады предоставить вам изображение под названием Винтовой датчик микрометра . Мы надеемся, что это изображение Микрометрический винтовой калибр поможет вам в изучении и исследовании. для получения дополнительных материалов по анатомии, подпишитесь на нас и посетите наш веб-сайт: www.anatomynote.com.
Анатомия.com нашел Микрометрическая структура винта калибра из множества анатомических изображений в Интернете. Мы думаем, что это наиболее полезный снимок анатомии, который вам нужен. Вы можете щелкнуть изображение, чтобы увеличить его, если не видите четкого изображения.
Изображение добавлено администратором. Спасибо за посещение anatomynote.com . Мы надеемся, что вы сможете получить именно ту информацию, которую ищете. Не забудьте поделиться этой страницей и подписаться на наши социальные сети, чтобы способствовать дальнейшему развитию нашего веб-сайта. Если у вас есть какие-либо вопросы, не стесняйтесь обращаться к нам.
Если вы считаете эту картинку полезной, пожалуйста, не забудьте поставить нам оценку под картинкой!
Одна из наших целей собрать эти изображения — мы надеемся, что эти изображения не будут потеряны при удалении соответствующей веб-страницы.
Но вы также можете знать, что любое содержимое, товарные знаки или другие материалы, которые могут быть найдены на веб-сайте anatomynote.com, которые не являются собственностью anatomynote.com, остаются собственностью соответствующих владельцев. Anatomynote.com никоим образом не претендует на право собственности или ответственности за такие предметы, и вам следует запросить юридическое согласие на любое использование таких материалов от его владельца.
Анатомия — удивительная наука. Это может помочь вам лучше понять наш мир. Мы надеемся, что вы будете использовать эту картинку в своем исследовании и в своих исследованиях.
Этот пост «Конструкция винтового датчика микрометра » относится к следующим категориям / категориям. В этих категориях вы также можете найти более похожее и подробное содержание.Физическая структура и микробный состав почвенных макроагрегатов в микрометрах
https: // doi.org / 10.1016 / j.soilbio.2013.02.005Получить права и контентAbstract
Макроагрегаты почвы представляют собой дискретные отдельные единицы почвы, которые, как мы предполагаем, содержат меньшие скопления микроорганизмов, чем массивная почва, и представляют масштаб, потенциально соответствующий естественным микробным сообществам . Мы задали два вопроса для изучения состава микробного сообщества в контексте макроагрегата: 1) Существует ли связь между физической структурой макроагрегата и составом микробного сообщества в отдельных макроагрегатах? И 2) Насколько похожи бактериальные сообщества в отдельных субмиллиметровых макроагрегатах почвы, взятых из одного и того же 5-сантиметрового керна? Для ответа на эти вопросы были взяты пробы отдельных макроагрегатов трех произвольных размеров (250–425, 425–841 и 841–1000 мкм) с поля пастбищ.Физические структуры 14 отдельных макроагрегатов были охарактеризованы с помощью просвечивающей рентгеновской томографии на основе синхротронного излучения, что показало, что большая часть порового пространства в макроагрегатах малого и среднего размера представляет собой поры относительно меньшего размера, что приводит к большей общей пористости и площадь границы раздела пор и минералов в этих более мелких макроагрегатах. Состав микробного сообщества охарактеризован с использованием данных пиросеквенирования 16S рРНК. Анализ разреженности показал, что членство в каждом макроагрегате было достаточно отобранным только с несколькими тысячами последовательностей; кроме того, членство в сообществах широко варьировалось между макроагрегатами, и структура варьировалась от сообществ, в которых сильно доминировало несколько филотипов, до сообществ, которые были равномерно распределены между несколькими филотипами.Мы не обнаружили прочной связи между физической структурой и членством в сообществе; это может быть связано с небольшим количеством агрегатов (10), по которым у нас есть как физические, так и биологические данные. Наши результаты подтверждают наше первоначальное ожидание, что отдельные макроагрегатные сообщества были значительно менее разнообразными, чем основная масса почвы с того же участка поля с пастбищами.
Основные моменты
► Изучены физическая структура и состав 16S отдельных макроагрегатов. ► Членство в сообществе было достаточно тщательно отобрано с <1000 последовательностей / совокупность.► Членство в сообществе и структура сильно различались между макроагрегатами. ► Просвечивающая рентгеновская микротомография показала большую пористость в более мелких макроагрегатах. ► Мы не обнаружили сильной корреляции между физической структурой и членством в сообществе.
Ключевые слова
16S
Макроагрегаты
Сообщества
Пиросеквенирование
Почва
Томография
Рекомендуемые статьи Цитирующие статьи (0)
Полный текстCopyright © 2013 Elsevier Ltd.Опубликовано Elsevier Ltd. Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
Новый сборщик метрик приложений Spring Boot 2
Micrometer — это фасад для сбора показателей , ориентированный на размерность , цель которого — позволить вам рассчитывать, подсчитывать и оценивать ваш код с помощью API, независимого от поставщика. С помощью пути к классам и конфигурации вы можете выбрать одну или несколько систем мониторинга для экспорта данных метрик. Думайте об этом как о SLF4J, но для показателей!
Микрометр— это средство сбора данных, включенное в привод Spring Boot 2.Он также был перенесен на Spring Boot 1.5, 1.4 и 1.3 с добавлением другой зависимости.
Micrometer добавляет более богатые примитивы счетчиков к счетчикам и датчикам, которые существовали в Spring Boot 1. Например, один таймер Micrometer Timer способен создавать временные ряды, связанные с пропускной способностью, общим временем, максимальной задержкой последних выборок, предварительно вычисленными процентилями , гистограммы процентилей и счетчики границ SLA.
Несмотря на то, что Micrometer сосредоточен на размерных показателях, он сопоставляет иерархические имена, чтобы продолжать обслуживать более старые решения для мониторинга, такие как Ganglia, или инструменты с более узкой областью действия, такие как JMX.Переход на Micrometer возник из-за желания лучше обслуживать волну систем размерного мониторинга (вспомните Prometheus, Datadog, Wavefront, SignalFx, Influx и т. Д.). Одна из сильных сторон Spring — это возможность выбора посредством абстракции. Благодаря интеграции с Micrometer, Spring Boot позволяет вам выбрать одну или несколько систем мониторинга для использования сегодня и изменить свое решение позже, когда ваши потребности изменятся, не требуя перезаписи инструментария пользовательских метрик.
Прежде чем выбрать «еще одну» библиотеку коллекции метрик, мы внимательно изучили существующие или перспективные сборщики измерений.Но по мере того, как мы рассматривали возможность экспорта во все больше и больше систем мониторинга, важность структуры имен и данных стала очевидной. Micrometer включает в себя концепцию нормализации соглашения об именах, масштабирования базовой единицы времени и поддержку частных выражений структур, таких как данные гистограммы, которые необходимы для того, чтобы метрики сияли в каждой целевой системе. Попутно мы также добавили фильтрацию счетчиков, что позволяет вам лучше контролировать инструментарий ваших зависимостей в восходящем направлении.
Анатомия пучков нервных волокон с микрометрическим разрешением в зрительной системе верветок
Резюме:
Такемура и его коллеги описывают подробный анализ траекторий основных путей белого вещества зрительной системы у верветок с помощью посмертной визуализации в поляризованном свете (PLI). Авторы четко очерчивают траектории как основных, так и второстепенных путей, обращаясь к разногласиям в литературе, с уровнем детализации, который еще предстоит сделать, учитывая ограничения предыдущих методов (например,g., диффузионная визуализация и диссекция Клингера). Исследование является исключительным с методологической точки зрения и очень подробным. Это феноменальный справочный документ, который будет иметь большое значение для нейроанатомии и визуальных наук.
Все три рецензента сочли это очень мощным и важным исследованием и поддержали его публикацию. Однако есть некоторые серьезные проблемы, которые необходимо решить, прежде чем он будет готов к принятию. Я хотел бы отметить, в частности, что озабоченность по поводу доступности данных (озабоченность, которую также разделяют рецензенты и старшие редакторы) особенно серьезна, учитывая приверженность eLife принципам открытой науки.
Мы благодарим редакторов и рецензентов за конструктивные комментарии и положительные оценки объема данной статьи. Мы обращаемся к каждому пункту ниже.
Существенные изменения:
1) Обрамление.
Рецензент №1 посчитал, что формулировка авторами мотивации этого исследования, направленная на устранение основных «разногласий» о существовании и траектории путей белого вещества в визуальном потоке, может преувеличить объем фактической работы. Во многих случаях такие споры об этих путях незначительны или давно отброшены в области нейроанатомии.Например, существование ILF подверглось сомнению в одном исследовании, проведенном еще в 1980-х годах. Большинство основных учебников нейроанатомии и области в целом признают его существование и соглашаются с основными траекториями ILF. Такое построение дает ложное представление о важности исследования. На самом деле, это исследование представляет собой анализ основных проводящих путей зрительной системы верветки, присутствующих у одних и тех же животных. Это достаточно существенный вклад в литературу без необходимости раздувать объем работы, поскольку она направлена на устранение критических разногласий, которые могут отсутствовать.
Благодарим рецензента за это предложение. Мы удалили тексты, относящиеся к «разногласиям» из аннотации и введения в измененной рукописи, и пояснили, что основная цель этого исследования — изучить детальную пространственную организацию волоконных трактов в зрительной системе верветок. Мы также согласны с рецензентами в том, что большинство исследователей больше не оспаривают существование ILF. Поэтому мы пересмотрели текст, заявив, что существование ILF было подвергнуто сомнению только в одном исследовании.
Мы также указали виды при описании литературы во Введении, следуя комментарию рецензента (см. Наш ответ на вопросы о видах ниже).
Конкретные изменения: В Резюме мы удалили тексты о противоречиях и реорганизовали тексты следующим образом:
«Хотя зрительная система приматов была тщательно изучена, детальная пространственная организация трактов волокон белого вещества, переносящих визуальную информацию между областями, не была полностью установлена.Это в основном связано с большим разрывом между исследованиями индикаторов и исследованиями диффузной МРТ, которые сосредоточены на конкретных аксональных связях и макроуровне организации трактов волокна, соответственно ».
Во Введении мы также внесли значительные изменения в абзац, первоначально относящийся к разногласиям:
«Несмотря на коллекции диссекционных, трассерных и дМРТ исследований зрительной системы, мы не до конца понимаем детальную пространственную организацию трактов белого вещества в зрительной системе, потому что остается большой разрыв между исследованиями, выполненными разными методами (Takemura et al. ., 2019b; Рашмор и др., 2020). В частности, хотя индикаторы хорошо подходят для измерения конкретных соединений от или к местам инъекции, этот метод не может визуализировать все тракты волокна. С другой стороны, хотя дМРТ хорошо подходит для измерения приблизительного положения и траекторий основных трактов волокон, у него недостаточно разрешения, чтобы точно измерить окончание трактов волокон в корковом сером веществе (Reveley et al., 2015). Таким образом, существует большой разрыв между данными о кортико-корковой связности, полученными в результате исследований с использованием индикаторов, и данными о трактах белого вещества, полученных в результате исследований dMRI.Более того, остается много нерешенных вопросов относительно пространственной организации трактов белого вещества, поскольку трудно точно измерить такую организацию с помощью любого из вышеупомянутых методов. Например, еще не ясно, насколько вертикальный затылочный пучок (VOF; Yeatman et al., 2014) является независимым пучком от нижнего продольного пучка (ILF) у макак (Schmahmann and Pandya, 2006; Takemura et al. , 2017). Более того, пространственная организация соседних трактов, таких как stratum sagittale (SS) и ILF, спорно обсуждается среди исследователей (Schmahmann and Pandya, 2006).Мы также отмечаем, что не во всех исследованиях сообщалось об одних и тех же трактах волокон или предлагалось идентичное определение трактов волокон (Schmahmann and Pandya, 2006; Yeatman et al., 2014). Эти неоднозначности в литературе частично являются результатом различных методов, используемых для каждого исследования (вскрытие, дМРТ и индикатор), поскольку эти методы имеют свои преимущества и ограничения. Исследование, визуализирующее целые пучки волокон с более высоким пространственным разрешением, кажется необходимым, чтобы заполнить пробел между различными методами и установить наше понимание детальной пространственной организации визуальных трактов белого вещества.”
В разделе Обсуждение мы написали:
«Хотя в прошлом существование ILF подвергалось сомнению (Tusa and Ungerleider, 1985), более поздние исследования трактографии на основе dMRI человека (Catani et al., 2003) и исследования индикаторов макак (Schmahmann and Pandya, 2006) продемонстрировали существование ILF как продольное связующее волокно ».
2) Ссылки на предыдущие работы.
У всех трех рецензентов возникли опасения по поводу связи исследования с предыдущей работой и существующими знаниями.
Рецензент№1 отмечает, что за последние 10 лет произошло значительное расширение анализа диффузионных изображений основных путей белого вещества, которые включают многие из тех же путей, изученных здесь (например, (Mori et al., 2008; Zhang) et al., 2008; Catani and de Schotten, 2008; Yeh et al., 2018)), включая фокальный анализ вентральных зрительных путей (например, (Pyles et al., 2013; Gschwind et al., 2012; Kamali et al., al., 2014; Toosy et al., 2004)). Чтобы оценить ценность текущей работы в перспективе, авторам необходимо провести более полное сравнение того, что известно в литературе, и сделать четкие различия в отношении видов.
Благодарим рецензента за указание на эти предыдущие работы. В отредактированной рукописи мы процитировали все эти предыдущие публикации и включили описания этих исследований dMRI в раздел «Введение» и «Обсуждение». Мы включили обсуждение литературы ILF в контексте категориальной обработки информации, цитируя работы, предложенные рецензентами, а также из других групп.
Мы также пересмотрели текст Введения, удалив наше описание противоречий и включив обширное обсуждение того, что известно, а что неизвестно в литературе по трассирующей и диффузионной МРТ.
Конкретные изменения: Во Вступление мы включили следующие тексты:
«Развитие методов получения и анализа dMRI привело к разработке атласов основных трактов волокон человека (Mori et al., 2008; Catani and Thiebaut de Schotten, 2008; 2012; Yeh et al., 2018) и автоматизированных процедур анализа эти участки основаны на данных dMRI (Zhang et al., 2008; Yendiki et al., 2011; Yeatman et al., 2012; 2018; Wassermann et al., 2016; Wasserthal et al., 2018; Warrington et al., 2020) ».
«Хотя эти участки уже были идентифицированы (исследования макак, Schmahmann and Pandya, 2006; Takemura et al., 2017; исследования на людях, Catani et al., 2003; Toosy et al., 2004; Catani and Thiebaut de Schotten, 2008). ; 2012; Kamali et al., 2014; Yeatman et al., 2014), эти исследования либо не предоставили прямых доказательств лежащей в основе анатомической структуры, либо подвержены методическим ограничениям (см. Обсуждение) ».
В разделе Обсуждение мы написали:
«Дальнейший анализ человеческого dMRI вместе с fMRI или поведенческими данными предполагает актуальность человеческого ILF с категориальной обработкой информации в вентральном визуальном потоке (Gschwind et al., 2012; Pyles et al., 2013; Scherf et al., 2014; Tavor et al., 2014) ».
В том же духе рецензент №2 отмечает, что выбор мозга верветы вызывает некоторое недоумение. Существует не так много предшествующей диффузной трактографии или анатомического прослеживания тракта, как на макаках-резусах. И, конечно, он несовершенный по сравнению с человеческим, и можно провести 3D-PLI на человеческом мозге (как, собственно, и сделали некоторые авторы). И горки не более близки к людям, чем макаки.Рецензент обеспокоен тем, что вместо того, чтобы прояснять, это еще больше загрязняет воду (и ограничивает воздействие) путем одновременного добавления нового вида и нового метода. К этому следует обратиться напрямую.
Как указал рецензент, в принципе, 3D-PLI можно применить к человеческому мозгу (например, Zeineh et al., 2017). На практике измерения 3D-PLI на человеческом мозге требуют значительно большего времени, чем измерения 3D-PLI на головном мозге приматов, кроме человека, потому что как количество, так и размер секций у людей значительно больше.Следовательно, сложно получить измерения на относительно большом количестве секций и нескольких мозгах. Мы решили провести измерения на двух мозгах верветок, чтобы сначала представить подробную структуру волокон модели животного, не являющегося человеком, приматов, в то время как мы продолжаем получать данные из человеческого мозга для будущих анализов. Кроме того, исследование 3D-PLI на приматах, отличных от человека, имеет преимущество перед исследованием 3D-PLI на людях в том смысле, что оно позволяет проводить прямое сравнение с богатой коллекцией измерений индикаторов и dMRI, опубликованных за последние несколько десятилетий (например,грамм. Шмахманн и Пандья, 2006; Schmahmann et al., 2007; Takemura et al., 2017).
Хотя рецензент отмечает, что наш выбор верветочной обезьяны может ограничить воздействие по сравнению с макаками, мы хотели бы поделиться своими взглядами на важность изучения верветочной обезьяны как модели приматов, не относящихся к человеку. Во-первых, мы хотели бы отметить, что, хотя мартышки-верветы не более тесно связаны с людьми, чем макаки, они не более отдаленно связаны с людьми, чем макаки. Таким образом, мы думаем, что разнообразие модельных видов приматов, не относящихся к человеку, поможет гарантировать, что выясненные тракты волокон более широко репрезентативны у обезьян старого мира, а не просто являются уникальной особенностью одного вида.Во-вторых, верветка становится все более важной моделью приматов, не относящихся к человеку, для исследований в области нейробиологии. Обезьяна верветки обычно изучалась в биомедицинских исследованиях в рамках более раннего обследования 2004 г. (Carlsson et al., 2004). Согласно опросу одного из авторов (N.P.G.), растет количество научных публикаций о верветке (изображение ответа автора 1). Эту тенденцию к росту можно также увидеть в отчете Национального института здравоохранения об оценке и анализе нечеловеческих приматов (2018; рисунок 5; https: // orip.nih.gov/sites/default/files/508%20NHP%20Evaluation%20and%20Analysis%20Final%20Report%20-%20Part%201.pdf). Недавний рост использования верветок может быть связан с тем, что они вызывают меньше опасений с точки зрения биобезопасности, поскольку они не являются носителями вируса герпеса B (Baulu et al., 2002), обычно дешевле, чем макаки (Fremier et al. , 2008) и появляющейся модели возрастных расстройств (Cramer et al., 2018; Latimer et al., 2019). Следуя этой тенденции, растет количество исследований нейробиологии, изучающих верветку (Fears et al., 2009; 2011; Woods et al., 2011; Федоров и др., 2011; Lundell et al., 2011; Dyrby et al., 2012; 2014; Maldjian et al., 2014; Menzel et al., 2019; Barrett et al., 2020). Подводя итог, мы хотели бы отметить, что (1) нет недостатка в изучении зрительной системы верветок с точки зрения эволюционного расстояния от людей и (2) становится все более распространенным изучение верветок в качестве модели приматов, не являющихся людьми. нейробиологические исследования.
Публикации PubMed о научных статьях, посвященных изучению верветок.
Вертикальная ось отображает количество публикаций за каждый год (с 1940 по 2019 год). Опрос проводился на сайте PubMed (https://www.ncbi.nlm.nih.gov/pubmed/) одним из авторов (N.P.G.).
Кроме того, мы также отмечаем, что теперь мы предоставляем доказательства, показывающие, что организация зрительной системы во многом схожа у мартышек вервет и макак. На изображении ответа автора 2 изображена закодированная цветом карта основного направления диффузии данных МРТ диффузии ex vivo, собранных из мозга верветок и макак.Хотя есть небольшие различия во взаимном расположении извилин и борозд, топологическое расположение пучков волокон, протестированных в этом исследовании (таких как SS, VOF и tapetum), согласовано между двумя видами. Мы также отмечаем, что детальная архитектура сливающихся волокон, окончаний волокон и U-волокон ближнего действия не видна в данных диффузионной МРТ. Поэтому мы думаем, что есть смысл выполнять измерения 3D-PLI с высоким разрешением на этих пучках в мозге верветок, и результаты, полученные по данным 3D-PLI верветок, могут быть переведены на макак, поскольку существует грубое межвидовое соответствие. пространственной организации волоконных трактов.Мы также отмечаем, что, основываясь на анализе данных авторадиографии рецепторов (Рисунок 12 — рисунок в приложении 1), парцелляция затылочной коры в целом сходна между мозгами макак и вервет. Таким образом, поскольку эти данные свидетельствуют о том, что зрительная система в значительной степени похожа у обоих видов с точки зрения как волоконных трактов, так и корковых областей, мы полагаем, что наш выбор верветки не ограничивает важность этой работы.
Сравнение затылочных участков белого вещества, видимых в данных диффузионной МРТ с цветовой кодировкой, между макаками и верветками.
( A ) Коронарное изображение кодированных с цветовой кодировкой данных диффузионной МРТ ex vivo, полученных от макак (левая панель; размер вокселя, изотропный 0,25 мм; 121 направление; b = 4800 с / мм 2 ; данные измерены в Национальном центре) Институты здравоохранения и предоставлены Д.А. Леопольдом и Ф.К. Й.) и мартышкой (правая панель; изотропный 0,5 мм; 128 направлений; b = 7700 с / мм 2 ; данные получены в Датском исследовательском центре магнитного резонанса и предоставлены Т. .B. Dyrby, следуя протоколам, описанным в Dyrby et al., 2011). Цветовая схема отображает главное направление диффузии в каждом вокселе (синий, верхний – нижний; зеленый, передний – задний; красный, левый – правый). Подробности этих наборов данных были описаны в предыдущих публикациях (данные о макаках, Thomas et al., 2014; Reveley et al., 2015; Takemura et al., 2017; данные Vervet, Donahue et al., 2016). ( B ) Осевой вид данных диффузионной МРТ ex vivo с цветовой кодировкой макак (левая панель) и верветок (правая панель).Несмотря на межвидовые различия в положении извилин и борозд и различия в параметрах сбора данных в наборах данных, положение stratum sagittale (SS), вертикального затылочного пучка (VOF) и тапетума (T) было согласованным между двумя видами.
Конкретные изменения: Мы включили параграф о полезности верхатной обезьяны в качестве модели приматов, не относящихся к человеку, в исправленную рукопись.
«Верветка как модель приматов, не относящихся к человеку, для нейробиологических исследований
В данном исследовании мы исследовали организацию волоконных трактов в зрительной системе верветок ( Chlorocebus aethiops sabaeus ).В то время как исторически макаки ( Macaca mulatta ) широко тестировались в исследованиях визуальной неврологии, верветки становились все более важной моделью для нейробиологических исследований из-за их биобезопасности (Baulu et al., 2002), более низкой стоимости (Fremier et al. , 2008) и сходство возрастных заболеваний с человеческими (Cramer et al., 2018; Latimer et al., 2019). Фактически, растет количество исследований в области нейробиологии, изучающих верветок в качестве модели приматов, отличных от человека, включая исследования, изучающие тракты волокон (Fears et al., 2009; 2011; Woods et al., 2011; Федоров и др., 2011; Lundell et al., 2011; Dyrby et al., 2013; 2014; Maldjian et al., 2014; Донахью и др., 2016; Menzel et al., 2019; Саруббо и др., 2019; Barrett et al., 2020). Мы также отмечаем, что, хотя мартышки-верветки не более тесно связаны с людьми, чем макаки, они не более отдаленно связаны с людьми, чем макаки. Мы думаем, что разнообразие модельных видов приматов, не относящихся к человеку, поможет гарантировать, что выясненные тракты волокон являются более широко репрезентативными для обезьян старого мира, а не просто уникальной особенностью одного вида.Кроме того, наши данные авторадиографии рецепторов также предполагают, что кортикальная область зрительной системы в целом схожа у мартышек и макак (рис. 12; рис. 12 — приложение к рисунку 1). Таким образом, настоящее исследование зрительной системы верветок с использованием 3D-PLI является важным шагом к пониманию организации зрительной системы приматов ».
Рецензент № 3 поднимает аналогичный вопрос, подчеркивая, что области зрительной коры идентифицируются со ссылкой на атласы, но они находятся у макак, а не у верветок.Это следует признать. Было бы неплохо увидеть немного больше обсуждения авторадиографии и критериев определения областей, поскольку это узкоспециализированные знания, которые мы должны принимать за чистую монету. По крайней мере, было бы полезно сослаться на различные критерии для определения областей.
Благодарим рецензента за предложения. В отредактированной рукописи мы дополнительно уточнили наши критерии для определения границы между корковыми областями на основе изображения пропускания данных 3D-PLI и авторадиографии рецепторов.Мы также дополнительно уточнили критерии, полученные на основе данных о верветах и макаках на каждом этапе. В отредактированной рукописи мы внесли значительные изменения в Методы, касающиеся идентификации корковых областей, а также включили подраздел, описывающий метод авторадиографии рецепторов. Наконец, мы отмечаем, что мы сделали данные 3D-PLI в этой работе общедоступными через платформу EBRAINS проекта Human Brain Project (DOI: 10.25493 / AFR3-KDK), чтобы предоставить данные, не зависящие от нашего определения границ (см. Наш ответ на комментарии к «Доступности данных» ниже).
Конкретные изменения: В разделе «Материалы и методы» написано:
«Количественная авторадиография рецепторов in vitro: экспериментальные процедуры, получение и обработка изображений
Каждый из замороженных блоков был серийно разрезан в корональной плоскости (толщиной 20 мкм) при -20 ° C с использованием микротома криостата (CM 3050, Leica, Германия). Срезы размораживали на предметных стеклах и сушили вымораживанием в течение ночи. Соседние срезы обрабатывали для гистологического окрашивания тел клеток (Merker, 1983) или миелиновых оболочек (Gallyas, 1979) или для количественной авторадиографии рецепторов in vitro в соответствии со стандартными протоколами (Palomero-Gallagher and Zilles, 2018; Zilles and Palomero-Gallagher, 2017). ).Глутаматергический каинатный рецептор, холинергический мускариновый рецептор M 2 и норадренергический рецептор α 1 визуализировали путем инкубирования соседних срезов в растворах соответствующих лигандов рецептора, меченного тритием. Неспецифическое связывание определяли в параллельных анализах связывания, в которых срезы инкубировали с меченным тритием лигандом и немеченым вытеснителем. Всем анализам связывания предшествовала предварительная инкубация в течение 20-30 минут в соответствующем буфере.
Каинатный рецептор метили [ 3 H] каинатом (8 нМ) в 50 мМ трис-цитратном буфере (pH 7.1), содержащий 10 мМ Ca-ацетата, при 4 ° C в течение 45 мин. Немеченый вытеснитель был каинатом (100 мкМ). За этой основной инкубацией следовала стадия промывки буфером (3 х 4 с) и два погружения в 100 мл ацетон, содержащий 2,5 мл глутаральдегида.
Холинергический мускариновый рецептор M 2 был помечен [ 3 H] оксотреморином-M (0,8 нМ) в 20 мМ буфере Hepes-Tris (pH 7,5), содержащем 10 мМ MgCl 2 при 22 ° C в течение 60 минут. мин. Немеченый вытеснитель представлял собой карбахол (1 мкМ).Основную инкубацию заканчивали стадией промывки буфером (2 х 2 мин) с последующим погружением в дистиллированную воду при 4 ° C.
Норадренергический рецептор α 1 метили [ 3 H] празозином (0,2 нМ) в 50 мМ трис-HCl буфере (pH 7,4) при 30 ° C в течение 45 минут. Немеченый вытеснитель представлял собой фентоламин (10 мкМ). Основную инкубацию заканчивали промывкой буфером (2 х 5 мин) с последующим погружением в дистиллированную воду при 4 ° C.
После анализа связывания срезы сушили в потоке теплого воздуха, затем совместно со стандартами известных концентраций радиоактивности подвергали воздействию тритий-чувствительных пленок (Hyperfilm, Amersham) для 4 (каинатный рецептор) или 6 (M 2 ). и α 1 рецепторов) недель.После проявления пленок авторадиографы были оцифрованы с помощью CCD-камеры (Axiocam MRm, Zeiss, Германия) и программного обеспечения для обработки изображений Axiovision (Zeiss, Германия). Кривая трансформации, показывающая взаимосвязь между значениями серого на авторадиографе и концентрациями радиоактивности в ткани, была рассчитана для каждого типа рецептора с использованием собственных сценариев Matlab (The MathWorks, Natrick, MA) и стандартов с известными концентрациями радиоактивности. Затем авторадиографы подвергали линейному усилению контраста, цветовому кодированию и медианной фильтрации, чтобы обеспечить четкую визуализацию региональных и ламинарных паттернов распределения рецепторов.”
Мы также писали:
«Идентификация корковых областей
Границы зрительных областей V1, V2, V3V, V4V, V4T, V5 / MT и MST, а также внутри теменных областей LIPe, LIPi, VIP и MIP могут быть идентифицированы у верветок на основе различий в их миелоархитектуре: проявляется как на изображениях пропускания, так и на картах ориентации волокон (Zilles et al., 2016). Эти области, а также области, определенные в теменной и височной долях, подтверждаются сравнениями между мозгом верветки 1818, обработанным для PLI, и мозгом верветки 1695, обработанным для гистологического окрашивания и авторадиографии рецепторов in vitro (Рисунок 12). .Хотя карты ориентации волокон и авторадиограммы были получены от разных животных, положение областей коры, идентифицированных различиями в миелоархитектуре, выявленных с помощью PLI (фиг. 12A), сравнимо с положением областей, выявленных с помощью паттернов распределения рецепторов (фиг. 12B-D). Наше разделение кортикальной ленты верветки также подтверждается параллелизмом между региональными и ламинарными паттернами распределения рецепторов передатчиков в мозге верветы и теми же рецепторами в срезах, полученных из сопоставимых ростро-каудальных уровней в мозге макака обезьяны (рис. Приложение к рисунку 1).Кроме того, области, идентифицированные в мозге макака, также были включены в атласы Салима и Логотетиса (2012 г.) и Паксиноса и др. (2009 г.).
Учитывая соответствие границ, идентифицированных анализом PLI и наборов авторадиографических данных рецепторов в головном мозге верветок, а также топологическую и архитектоническую сопоставимость рецепторов областей, идентифицированных в головном мозге верветок и макак, разделенных на коронку, области коры в сагиттально разрезанном головном мозге верветки (ID 1947) были идентифицированы на основе региональных и ламинарных различий в распределении рецепторов передатчиков в сагиттальных срезах мозга макаки (Zilles and Clarke, 1997), а также путем сравнения с атласами Saleem и Logothetis (2012) и Paxinos et al. ., (2009).
Мы сделали данные 3D-PLI, используемые для генерации цифр, общедоступными через платформу EBRAINS проекта Human Brain Project, чтобы предоставить данные, объективные по нашим определениям границ. Данные (Axer et al., 2020) доступны через следующий DOI: 10.25493 / AFR3-KDK ».
3) Краткое изложение работы.
Рецензенты №1 и №2 отмечают, что авторы проделали фантастическую работу по характеристике этих путей с разных точек зрения. И все же пресловутый лес теряется за деревьями (т.е. сложно отслеживать детали изображений и текстовых описаний). Оба рецензента считают, что визуальное резюме траектории всех основных путей, обозначенных здесь, будет иметь большое значение для помощи читателям в понимании главной идеи статьи.
Следуя предложению рецензента, мы включили существующую литературу, а также новый рисунок, выделяющий предыдущие работы по рассечению, трассирующей или диффузионной МРТ, чтобы предоставить читателям обзор приблизительных положений и траекторий основных трактов в зрительной системе приматов (рис. рукопись).Мы также включили Таблицу 1 в исправленную рукопись с описанием траектории основных путей (см. Наш ответ рецензенту № 2 ниже). Мы считаем, что результаты нашего настоящего исследования улучшают наше понимание этих участков, а на Рисунке 1 подчеркивается необходимость подхода с высоким разрешением, использованного в этом исследовании, чтобы точно понять пространственную организацию этих участков.
В разделе результатов мы написали.
«На рисунке 1 изображены описания основных трактов в зрительной системе приматов, stratum sagittale, нижнего продольного пучка (ILF) и вертикального затылочного пучка (VOF), как определено в предыдущих исследованиях диссекции на людях (рис. 1A-B), и макак tracer (рис. 1C) и dMRI (рис. 1D) исследования.Хотя эти исследования выявили приблизительное положение и траекторию каждого тракта (определение этих трактов см. В Таблице 1), ни одно из них не выявило пространственную организацию всего рассматриваемого тракта отдельного волокна с микрометровым разрешением. Поэтому мы стремились исследовать детальную пространственную организацию этих участков и других участков, не описанных на рисунке 1, с использованием подхода 3D-PLI на визуальной системе верветок ».
Рецензент№2 предлагает нечто подобное трехмерным изображениям из их статьи 2016 года, а также некоторые черно-белые корональные срезы в стиле атласа, показывающие границы различных связок.Этот рецензент указал, что есть ряд потенциальных читателей, которые не захотят углубляться в цветовые схемы, используемые на текущих рисунках, но захотят узнать расположение пучков и общую ориентацию волокон.
Мы понимаем, что многие читатели привыкли видеть схему цветового кодирования RGB, широко используемую в клинических исследованиях диффузной МРТ (например, Pajevic and Pierpaoli, 1999). Однако мы часто обнаруживаем, что эта схема визуализации не полностью захватывает информацию о данных 3D-PLI, поскольку она присваивает идентичные цвета волокнам с ориентацией, которая является зеркально-симметричной по отношению к вертикальному меридиану.Поэтому мы думаем, что цветовая схема RGB часто теряет важную информацию в данных 3D-PLI с высоким разрешением. По этой причине цветовое кодирование HSV часто использовалось для исследований PLI, даже когда исследование было направлено на сравнение PLI и dMRI (Henssen et al., 2019a; 2019b). Мы включаем изображение ответа автора 3 для справки по сравнению обеих схем цветового кодирования, а также включаем ссылку на предыдущую работу по сравнению схем цветового кодирования (Axer et al., 2011; Henssen et al., 2019a; 2019b) в переработанная рукопись.
Мы также понимаем, что будет гораздо предпочтительнее получить точные оценки трехмерной функции распределения ориентации и адаптировать их к визуализациям. Однако такая оценка требует расширения поляризованной микроскопии косыми измерениями (Schmitz et al., 2018). Эффективный протокол для таких измерений все еще находится в стадии разработки. См. Наш ответ на комментарии об ограничении 3D-PLI ниже.
Сравнение схем кодирования цветов для данных 3D-PLI.
( A ) Цветовое представление HSV данных 3D-PLI, полученных из медиального сагиттального разреза через левое полушарие мозга верветок (ID1947; раздел № 249). Изображение идентично изображенному на рисунке 2А. ( B ) Цветовое представление данных 3D-PLI в формате RGB. Раздел идентичен разделу, представленному на панели A. Сфера указывает цветовую кодировку RGB ориентации трехмерного волокна в каждом пикселе на панели B.
Конкретные изменения: В разделе «Материалы и методы» написано:
«Сравнение с другими схемами кодирования цвета, часто используемыми в исследованиях dMRI, было описано в предыдущих публикациях (Axer et al., 2011b; Henssen et al., 2019a; 2019b)».
В разделе Обсуждение мы написали:
«Мы суммируем наши наблюдения в Таблице 1, при этом отметив, что все кортикальные окончания каждого пучка могут не быть включены в эту таблицу, поскольку некоторые из них все еще трудно идентифицировать из-за пересечения с поверхностными U-волокнами или слияния с другими трактами.”
Рецензент№2 отмечает, что дополнительная таблица, суммирующая точки входа / выхода для каждого из различных пучков волокон, была бы очень полезной. Одним из преимуществ этого метода перед диффузионной трактографией является улучшенная способность различать их — при разрешении 1,3 мкм 3D-PLI приближается к ширине одиночного аксона. Хорошо, что некоторые из них неясны из-за пересечения U-волокон, которые могут быть включены в таблицу.
Благодарим рецензента за это предложение.В раздел «Обсуждение» мы включили Таблицу 1, которая представляет собой сводку описаний. Обратите внимание, что, как упомянул рецензент, невозможно включить все кортикальные окончания волоконного тракта.
Конкретные изменения: Добавлена таблица 1.
В разделе Обсуждение мы написали:
«Мы суммируем наши наблюдения в Таблице 1, при этом отметив, что все кортикальные окончания каждого пучка могут не быть включены в эту таблицу, поскольку некоторые из них все еще трудно идентифицировать из-за пересечения с поверхностными U-волокнами или слияния с другими трактами.”
4) Четкость выводов.
Рецензент №2 спрашивает, насколько уверены авторы в том, что VOF на самом деле в основном переносит волокна от дорсальных к вентральным областям мозга, а не переносит дорсальные и вентральные волокна к середине, чтобы пересечь рострально / каудально (скажем, в ILF?).
Как в корональном, так и в сагиттальном срезе, мы наблюдали, что волокна VOF соединяют дорсальную и вентральную экстрастриатную кору, и мы не наблюдали аксонов с резкими поворотами и оканчивающимися медиально.Поскольку ложноотрицательные результаты вполне возможны при любом экспериментальном подходе, мы допускаем, что мы не можем исключить возможность того, что небольшое количество аксонов покидает VOF и принимает рострально-каудальное направление. В исправленную рукопись мы включили обсуждение этого вопроса.
Конкретные изменения: В разделе «Обсуждение» мы написали:
«Хотя мы не можем исключить возможность того, что небольшое количество аксонов может покинуть VOF и принять ростро-каудальное направление, наши наблюдения в данных 3D-PLI подтверждают точку зрения, что волокна VOF в основном перемещаются вдоль верхней-нижней оси и соединяют дорсальную и вентральную экстрастриатную кору.”
Рецензент №3отмечает, что авторы могли бы рассмотреть возможность предоставления немного большего контекста в разделе «Результаты». Многие трактаты обсуждаются без какой-либо предыстории, и логика их решения называть бандл определенным именем остается неявной. Несомненно, у авторов есть веские причины для обозначения своих трактатов, поэтому было бы неплохо, если бы некоторые из этого неявного — и, без сомнения, огромного — хранилища знаний были разделены. В разделе «Обсуждение» есть обширный обзор, но раздел «Результаты» можно было бы сделать более читабельным.В общем, более обширная ссылка на литературу принесет пользу статье. Например, неоднократно упоминается «полемика» вокруг VOF, но это подтверждается ссылкой на статью, в которой идентифицирован трактат, — вряд ли это утверждение противоречия.
В ответ на комментарий рецензента и другие комментарии мы внесли несколько изменений, чтобы помочь читателям понять основу волоконных трактов в зрительной системе приматов. Во-первых, мы включили рисунок в раздел «Результаты», изображающий предыдущую литературу по основным трактатам для ознакомления читателей (рисунок 1; см. Наш ответ на комментарий рецензента №1 и №2 «Краткое изложение работы» выше).Во-вторых, мы включаем таблицу с описанием имени, местоположения и траектории, а также корковых окончаний, наблюдаемых в этом исследовании, в виде таблицы 1 (см. Наш ответ на комментарии рецензента №2 в таблице). Таблица 1 также суммирует определение каждого тракта вместе со ссылками на предыдущие работы, в которых установлено данное определение.
Мы также отмечаем, что мы удалили тексты, относящиеся к «разногласиям», следуя предложениям рецензента №1 (см. Наши ответы рецензенту №1).
Наконец, в процессе редактирования мы заметили, что рукопись не содержала обширных ссылок на предыдущие работы над тапетумом.Теперь мы включили абзацы в раздел «Обсуждение» тапетума со ссылкой на предыдущие работы.
Конкретные изменения: Рисунок 1 и Таблица 1.
В разделе «Обсуждение» мы написали:
“ Tapetum
В то время как ряд нейроанатомов в XIX веке сообщали о существовании тапетума, существовала значительная путаница относительно того, следует ли рассматривать этот пучок волокон как ассоциативное волокно или как мозолистое волокно (см. Schmahmann and Pandya, 2006; 2007; Forkel et al. al., 2015 за исторические дебаты о тапетуме). Среди классических нейроанатомов Burdach (1822) сообщил, что волокна тапетума являются продолжением мозолистого тела splenium corpus. Поскольку это наблюдение было позже подтверждено рядом исследователей, использовавших различные методы, мозолистое происхождение волокон тапетума больше не может обсуждаться (Mettler, 1935; Clarke and Miklossy, 1990; см. Обзор Schmahmann and Pandya, 2006). Данные 3D-PLI действительно непосредственно визуализировали, что волокна тапетума переходят в мозолистое тело I (рис. 11), подтверждая эти предыдущие работы.Данные 3D-PLI дополнительно продемонстрировали подробный ход волокон тапетума, а именно его относительный ход по отношению к соседним пучкам (SS и stratum calcarinum, как обсуждается ниже), боковому желудочку и эпендиме (рисунки 9C, 10C, 11 и рисунок 9 — дополнение к рисунку) 1). Эта подробная информация предоставит важную информацию для руководства трактографическими исследованиями на основе dMRI на волокнах селезенки, которые считаются важными для важных корковых функций, таких как чтение (Binder and Mohr, 1992; Dougherty et al., 2007) ».
5) Доступность данных.
Рецензенты № 1 и 3, а также старший редактор считают, что доступность данных, предоставляемая авторами, не соответствует стандартам, требуемым eLife. Рецензент №3 отмечает, что авторы заявляют, что все данные представлены на рисунках, но, конечно, это даже близко не демонстрирует богатство их данных. В эту эпоху открытой науки мы очень надеемся, что они создадут ресурс для ученых, основанный на их данных; это также был бы единственный способ, которым читатель может действительно оценить свои утверждения, поскольку, очевидно, текущая рукопись по своей природе является описательной и зависит от разделов, которые авторы выбирают для показа.Рецензент понимает, что эти данные, вероятно, настолько велики, что делают простой обмен в Интернете запретительным, но авторы будут приветствовать определение траектории для обмена данными такого типа. По крайней мере, им следует перефразировать свое заявление о доступности данных.
Благодарим редакцию и рецензентов за это предложение. Как предполагают обозреватели, учитывая большой размер файла данных 3D-PLI, для нас нецелесообразно использовать широко используемые общедоступные онлайн-репозитории.Создание инфраструктуры для обмена такими большими данными — это постоянная работа в рамках проекта Human Brain Project (HBP). В ответ на комментарии мы делаем данные 3D-PLI, проанализированные в этой рукописи, общедоступными через платформу EBRAINS проекта Human Brain Project (DOI: 10.25493 / AFR3-KDK). Кроме того, мы включили адрес электронной почты одного из авторов-корреспондентов в Исследовательском центре Юлих (N.P.G.), чтобы предоставить контактную информацию для ученых, которые могут пожелать получить доступ к данным, еще не предоставленным на платформе HBP.
Конкретные изменения: В заявлении о доступности данных мы написали:
«Исходные данные общедоступны через платформу EBRAINS проекта Human Brain Project (Axer et al., 2020; DOI: 10.25493 / AFR3-KDK)».
В разделе «Материалы и методы» мы писали:
«Мы сделали данные 3D-PLI, используемые для создания цифр, общедоступными через платформу EBRAINS проекта Human Brain Project, чтобы предоставить данные, объективные по нашим определениям границ. Данные (Axer et al., 2020) доступен через следующий DOI: 10.25493 / AFR3-KDK ».
https://doi.org/10.7554/eLife.55444.sa2Структуры MicroED из микрометрических толстых кристаллов белка
Abstract
Теоретические расчеты показывают, что кристаллы толщиной более 100 нм исключаются из-за динамического рассеяния из успешного определения структуры с помощью дифракции электронов на микрокристаллах (MicroED). Эти расчеты расходятся с экспериментальными результатами, в которых структуры MicroED были определены из значительно более толстых кристаллов.Здесь мы систематически оцениваем влияние толщины на точность интенсивности MicroED и способность определять структуры из кристаллов белка толщиной в один микрометр. Для этого мы сравниваем структуры ab initio сегмента прионного белка человека, определенные на основе тонких кристаллов, с структурами, определенными для кристаллов толщиной до одного микрометра. Мы также сравниваем растворы для замены молекул из кристаллов различной толщины для более крупного глобулярного протеина, протеиназы K. Наши результаты показывают, что структуры могут быть надежно определены из кристаллов, по крайней мере, на порядок более толстых, чем предполагалось ранее при моделировании, что открывает возможность даже для более широкий спектр экспериментов MicroED.
Резюме С помощью MicroED можно определить белковые структуры с атомным разрешением на кристаллах, которые на порядок превышают теоретический предел максимальной толщины.
Основной текст
Введение
Электроны взаимодействуют с веществом сильнее, чем рентгеновские лучи, и предлагают большую долю полезных событий упругого рассеяния по сравнению с событиями неупругого рассеяния (1). Эти свойства используются методом криоЭМ, MicroED, для определения структуры с атомным разрешением из наноразмерных кристаллов белка (2).С помощью этого метода дифракция измеряется от белковых нанокристаллов в замороженном-гидратированном состоянии (3) с использованием пучка электронов с низкой дозой (обычно ~ 0,01 e — Å -2 s -1 ) (4–6). Кристаллы непрерывно и однонаправленно вращаются в луче, в то время как дифракционные изображения собираются в виде фильма на быстром детекторе (7). Ряд ранее неизвестных, а также известных структур был определен этим методом с атомным разрешением (8).
Сильное взаимодействие между электронами и материалами также позволяет иметь место многократному рассеянию до того, как электроны покинут образец (9).Согласно теории динамического рассеяния (10), события многократного рассеяния приводят к неточностям в регистрируемых отражениях, потенциально препятствуя растворению структур. Моделирование показывает, что для кристаллов толщиной более 50-100 нм (11, 12) динамическое рассеяние может быть сильным, что приводит к почти случайным интенсивностям, когда связь между интенсивностью и структурным фактором больше не выполняется (10). Однако это моделирование предполагает, что дифракция регистрируется от идеального и неподвижного кристалла — настоящие макромолекулярные кристаллы не идеальны.Фактически, прецессионная дифракция электронов позволяет избежать многих артефактов, связанных с динамическим рассеянием от почти идеальных кристаллов неорганического материала, за счет поворота электронного луча вокруг кристалла (13). Точно так же, используя непрерывное вращение MicroED (7), полезная дифракция обычно снимается с кристаллов белка толщиной в сотни нанометров (14). Было показано, что даже кристаллы лизоцима толщиной 1,5 мкм дают дифракцию, которая при объединении дает разумную статистику (2).Более того, недавние структуры, определенные с помощью методов ab initio с разрешением 1 Å, дополнительно указывают на то, что интенсивности дифракции, полученные при непрерывном вращении MicroED, являются точными и поддерживают соотношение между амплитудой и фазой (4, 15).
Здесь мы систематически исследуем взаимосвязь между толщиной кристалла, динамическим рассеянием и качеством структурных решений, полученных с помощью MicroED. Мы определяем ab initio структур сегмента из петли β 2 -α 2 человеческого прионного белка в амилоидном состоянии, а также структуру протеиназы K путем молекулярного замещения кристаллов толщиной до микрометра.
Результаты
Сравнение структур, определенных из тонких и толстых кристаллов сегмента β2-α2 петли человеческого прионного белка
В качестве модельной системы для оценки влияния толщины кристаллов на интенсивность дифракции мы используем сегмент из петля β2-α2 человеческого прионного белка (hPrP), которая содержит остаток глицина на своем аминоконце (последовательность GSNQNNF), далее именуемую hPrP-β2α2, для анализа структуры MicroED из кристаллов различной толщины.Кристаллы этого сегмента выглядят как иглы длиной в микрометр и имеют толщину от нескольких нанометров до более микрометра. Были собраны семь наборов данных, полученных из тонких кристаллов, и объединены для получения набора справочных данных, который был на 80,3% завершен в P1 с константами { a , b , c } (Å) = {4.86, 14.11, 18.41} , и углы {α, β, γ} (°) = {90,00, 93,71, 101,21}. Второй набор данных был построен из 7 кристаллов толщиной от 500 нанометров до одного микрометра, которые были объединены в 75.Полный набор данных на 6% с той же пространственной группой и размерами элементарной ячейки, как указано выше. Тонкие и толстые кристаллические структуры были определены ab initio прямыми методами с использованием SHELXT (16) (см. SI Methods ) и уточнены до атомного разрешения с помощью BUSTER-TNT (17), показав четкую атомную плотность ( Рисунок 1) . Статистика уточнения для hPrP-β2α2 представлена в таблице 1 .
Рисунок-1: Структурное сравнение hPrP-β2α2 из тонких и толстых кристаллов прямыми методами( A ) Структурная модель для hPrP-β2α2, полученная из кристаллов размером <200 нм ( B ) из набора кристаллов со средним толщина> 1 мкм.Карты плотности 2mFo-DFc и mFoDFc очерчены на уровнях 1,5 σ и 3 σ соответственно. Положительные плотности водорода для обоих растворов показаны в A и B зеленым цветом и обозначены оранжевыми стрелками. ( C ) Растворы Ab initio как из нормальных (желтый), так и из кристаллов пептида размером> 1 мкм (синий). ( D ) Окончательная структурная модель hPrP-β2α2.
Таблица-1:Статистика сбора и уточнения данных.
Структура hPrP-β2α2 представляет собой прионный протофиламент с амилоидными свойствами.Протофиламент представляет собой стерическую молнию класса 2, состоящую из параллельных бета-листов лицевой стороной к спине (18). Одна пара листов образует протофиламент, как и другие амилоидные структуры (15, 18). При таком разрешении плотность показывает атомы водорода, а также присутствие ионов цинка и ацетата, которые облегчают кристаллографические контакты; оба присутствовали в состоянии кристаллизации ( Рисунок 1 ). Структуры как из толстых, так и из тонких кристаллов показывают четкую плотность воды на карте 2F o -F c и имеют несколько водородов, появляющихся на карте F o -F c на уровне 3 σ ( Рисунок 1 ).О появлении разрешимых водородов при дифракции электронов впервые сообщили для белков Rodriguez et al. (15), а затем Палатинусом (19) для малых молекул. Наши растворы ab initio для тонких и толстых кристаллов очень похожи, со среднеквадратичным отклонением основной цепи 0,07 Å и среднеквадратичным отклонением для всех атомов 0,09 Å ( Рисунок 1) . Таким образом, мы не обнаружили существенных различий между структурами, определенными из тонких и толстых кристаллов hPrP-β2α2.
Сравнение структур, определенных из тонких и толстых кристаллов глобулярного белка
Мы собрали данные MicroED для одиночных кристаллов протеиназы K толщиной ~ 500 нм и ~ 1 мкм ( SI Рисунок 2, SI Рисунок 3 ) и определили конструкции из каждого.Отражения регистрировались с разрешением 1,8 Å в обоих случаях, что дает полноту 97% для кристалла 500 нм и полноту 79,7% для кристалла толщиной 1 мкм. Данные были сокращены в XDS (20) и поэтапно заменены молекулами с использованием атомных координат PDBID 5i9s в качестве модели поиска (21). Структуры уточняли с помощью phenix.refine (22) с отсечкой высокого разрешения 2,0 Å и 3,0 Å соответственно (, таблица 1, ; , рисунок 2, ). Поскольку единственная кристаллическая структура толщиной ~ 1 микрометр имела плохую статистику, ее данные были объединены с данными, полученными от трех дополнительных кристаллов протеиназы K; каждый из них также имеет толщину ~ 1 мкм ( SI Рисунок 4, SI Рисунок 5 ).Структура из четырех объединенных наборов данных была снова определена с использованием той же модели поиска теперь с разрешением 2,5 Å с лучшей общей статистикой ( Таблица 1 , Рисунок 2 ). Сравнение полученных структур с моделью поиска молекулярного замещения показывает хорошее согласие со среднеквадратичным отклонением всех атомов ниже 0,25 Å. Таким образом, мы не обнаружили существенных различий между структурами, определенными из тонких и толстых кристаллов протеиназы K.
Рисунок 2: Сравнение структур глобулярных белков, определенных из тонких, промежуточных и толстых кристаллов.Плотность боковой цепи протеиназы К и соответствующий структурный раствор из ( A ) тонких кристаллов размером ~ 200 нм (PDBID 5I9S) (20), ( B ) одного кристалла толщиной 600 нм, ( C ) одного кристалла 1000 нм- толщиной кристалл и ( D ) четыре кристалла толщиной более 1000 нм. Все карты плотности 2F o -F c имеют контур на уровне 1,5σ для остатков 226-240, показанных серой сеткой. ( E ) Все структурные растворы протеиназы К наложены для сравнения.Наложенные цвета соответствуют индивидуальным цветным структурам.
Систематическое исследование влияния толщины кристалла на качество данных MicroED
Дифракция была измерена на 19 кристаллах hPrP-β2α2 с толщиной от ~ 100 нм до ~ 1100 нм на клиньях аналогичного размера обратного пространства, соответствующих реальному космический угловой диапазон приблизительно от -30 ° до + 30 ° ( SI Таблица 1 ). Данные для всех сравнений были проиндексированы и интегрированы с помощью XDS (20). Оценка толщины кристалла подробно обсуждается в SI Materials .Вкратце, средняя толщина кристалла оценивается путем измерения площади проецирования по изображениям, записанным при наклоне 0 ° и 60 °, а соотношение сторон соответствует идеализированной модели. Эти геометрические оценки были подтверждены отношениями интенсивности с использованием подсчета кадров, как описано ранее (14, 23), и хорошо согласуются ( Si, рис. 6, ). Изображения кристаллов представлены в документе SI . Чтобы оценить качество дифракционных данных, мы сравниваем значения R (Rmeas), отношение интенсивности к дисперсии (I / σI) и коэффициент корреляции половинного набора (CC1 / 2).Все измеренные значения данных и статистические данные представлены в SI Таблица 1 . Измеренные значения R для этих 19 кристаллов показывают среднее значение 13,03% со стандартным отклонением 2,4%; Среднее значение I / σI составляет 4,1 со стандартным отклонением 0,91. Коэффициент корреляции половинного набора составляет в среднем 98,2% со стандартным отклонением 1,4%. Амплитуды структурных факторов для этих 19 кристаллов сравнивали со структурным раствором hPrP-β2α2 из тонких кристаллов, обсуждаемых выше. Коэффициент корреляции между решенной моделью и отдельными кристаллами показан на Figure-2D как CC model ( Figure 3; SI Table 1 ).
Рисунок-3: Измерения на кристаллах hPrP-β2α2 переменной толщины.( A ) Средние значения I / σI, ( B ) Измеренные, ожидаемые значения R и их разность, Rdiff, ( C ) внутренний коэффициент корреляции половинного набора и ( D ) корреляция измеренных структурные факторы к решенной модели тонкого кристалла, или решенная модель CC . Средние значения показаны сплошными линиями. Пунктирная черная линия соответствует максимальному зарегистрированному пределу моделирования (10).
Поглощение кристаллами белка
Данные, представленные выше, показывают, что динамическое рассеяние не препятствует растворению структуры с помощью MicroED даже из кристаллов микрометровой толщины, когда данные собираются путем непрерывного вращения. Однако следует отметить, что достижимое разрешение было ниже для толстых кристаллов по сравнению с тонкими кристаллами (Рисунок 1,2 и Таблица 1) . Чтобы оценить, ограничивает ли поглощение толстыми кристаллами достижимое разрешение, мы записали спектры электронной спектроскопии потерь энергии (EELS) для 11 кристаллов hPrP-β2α2 и 10 кристаллов протеиназы K с толщиной от ~ 100 нм до ~ 1400 нм при 300 кВ ( Рисунок 4 ).Контрольные спектры записывали из областей пустой углеродной подложки или внутри отверстий решетки ( SI Document 3 ). Каждое изображение было выровнено с помощью анализа главных компонентов, и линейное сканирование через пик с нулевыми потерями было измерено вдоль первого главного компонента. Интенсивности пиков с нулевыми потерями показывают ожидаемый экспоненциальный спад (9). Существенная потеря энергии наблюдалась только для углеродной подложки с уменьшением интенсивности прошедшего луча более чем на 40% ( SI Document 3 ). Экспоненциальная аппроксимация данных (, рис. 4, ) предполагает, что длина затухания или (1 / e) потеря интенсивности из-за углеродной пленки будет ~ 87 нм при 300 кВ.Однако потери (1 / e) кристаллов протеиназы K и hPrP-β2α2 составляют 261 нм и 323 нм соответственно при 300 кВ. Наши оценки фактической толщины углерода составляют около 45 нм с использованием соотношений интенсивностей ( SI, рис. 6, ), что хорошо согласуется с предыдущими выводами по этим сеткам (24). Таким образом, даже с учетом подложки из углеродной пленки наши оценки очень близки к средней длине свободного пробега воды при 300 кВ, которая экспериментально составляет 336 нм (25, 26). Эти данные свидетельствуют о том, что даже трехкратного уменьшения интенсивности из-за поглощения недостаточно, чтобы запретить решение структурных решений с помощью MicroED для макромолекулярных кристаллов микромолекулярной толщины при 300 кВ; достижимое разрешение быстро падает вблизи этого предела толщины или за его пределами.
Рисунок 4: Наблюдаемое пропускание кристаллов разной толщины.Пиковые интенсивности спектров электронной спектроскопии потерь энергии (EELS), полученные для кристаллов hPrP-β2α2 (синий) и протеиназы K (оранжевый). Черные точки соответствуют зарегистрированной интенсивности вакуума и углеродной пленки. Модели экспоненциального затухания представлены пунктирными линиями, согласованными с цветом образца. Предел толщины для экспериментов MicroED, предложенный simulatins, показан штриховой горизонтальной линией (10).
Обсуждение
Эти данные показывают, что с помощью MicroED можно собрать точные данные и надежно определить структуры из кристаллов, которые на порядок толще, чем предполагалось ранее. Моделирование показало, что определение структуры с помощью MicroED будет тормозиться динамическим рассеянием кристаллов толщиной всего ~ 50-100 нм (11, 27). Здесь мы показываем, что растворы белковой структуры могут быть получены из кристаллов значительно большей толщины, чем предполагают смоделированные пределы как для глобулярных белков, так и для пептидов, с использованием методов молекулярной замены и ab initio соответственно.
Структурные решения высокого качества могут быть получены как для hPrP-β2α2, так и для протеиназы К независимо от данных, полученных от тонких кристаллов или кристаллов толщиной около одного микрометра. Четкая атомная плотность на карте 2F o -F c hPrP-β2α2 выявляет атомы водорода и присутствие цинковых и ацетатных лигандов, а также упорядоченных молекул воды. Множественные водороды проявляются в плотности F o -F c на уровне 3 σ (, фиг. 1, ). Точно так же плотность протеиназы K имеет высокое качество с четко определенной плотностью как на σ-взвешенных разностных картах, так и на композитных картах пропусков SA для данных, полученных из тонких (<200 нм), промежуточных (~ 500 нм) и толстых (> 1000 нм). ) кристаллы.Толщина не препятствует сбору, интеграции или определению структуры данных из этих кристаллов.
Исходный структурный раствор протеиназы К, определенный с помощью MicroED ( 5i9s ), был уточнен до разрешения 1,75 Å по объединенным данным четырех тонких кристаллов (20). Наши настоящие модели из монокристаллов толщиной ~ 500 нм и ~ 1000 нм определены с разрешением 2 Å и 3Å соответственно. Мы связываем эту разницу в разрешении с увеличением поглощения, что продемонстрировано нашим анализом EELS для более толстых кристаллов.Вместе эти результаты показывают, что достижимое разрешение может быть ограничено толстыми образцами. Однако объединение данных из нескольких толстых кристаллов дало улучшенную статистику и, следовательно, улучшенное разрешение, что указывает на то, что явления поглощения можно до некоторой степени преодолеть за счет увеличения избыточности измерений. Разрешение более толстых кристаллов протеина в конечном итоге ограничивается абсорбцией.
Почему существует несоответствие между экспериментальными данными, представленными здесь, и пределами, найденными в предыдущих моделях? Сложность, присущая моделированию, требует допущений об условиях эксперимента, которые не отражают сложность экспериментов MicroED.В частности, при моделировании предполагается, что кристаллы идеальны, все электроны рассеяны, а данные записываются с неподвижных кристаллов от осей основных зон (11, 28). Эти условия часто встречаются в кристаллах небольших молекул, таких как неорганические соединения в материаловедении, и в двумерных (2D) кристаллах белка, но редко встречаются в кристаллографии макромолекул с использованием 3D-кристаллов белка и протоколов MicroED (29). Кристаллы макромолекул сильно мозаичны по сравнению с кристаллами простых органических соединений или неорганических кристаллов.Макромолекулярные кристаллы несовершенны и искривлены в наномасштабе. Данные MicroED от высокомозаичных кристаллов выигрывают от их беспорядка по аналогии с поворотом электронного луча при прецессионной дифракции электронов. Кроме того, данные MicroED собираются путем непрерывного вращения, поэтому интеграция по кривой качания дополнительно ограничивает динамические эффекты. Наши результаты подтверждают, что динамическое рассеяние не является основной проблемой при решении кристаллических структур белка, и что эффекты динамического рассеяния не увеличиваются линейно с толщиной кристалла.Однако вопрос о том, почему не наблюдаются значительные артефакты из-за многократного рассеяния, все еще остается и требует дальнейшего изучения.
Тот факт, что артефакты множественного рассеяния не ограничивают определение структуры в MicroED, особенно удивителен для кристаллов толщиной более ~ 300 нм, где толщина кристаллов легко превышает длину свободного пробега электрона при 200 или 300 кВ ( Рисунок 4 ) ( 25, 26, 30). Для этих кристаллов через кристалл проходит небольшое количество электронов без какого-либо взаимодействия с образцом.При очень больших толщинах дифракционные пятна начинают расширяться из-за потерь энергии. Более крупные пятна затрудняют измерение кристаллов с большим количеством атомов на элементарную ячейку и могут дать плохую статистику, поскольку большая часть рассеяния будет уменьшена за счет поглощения. Объединение больших уширенных пятен кристаллов с крупными элементарными ячейками может привести к перекрытию пятен и другим осложнениям, которые могут ограничить достижимое разрешение; мы еще не сталкивались с этой проблемой. Самым крупным белком, определенным на сегодняшний день с помощью MicroED, является каталаза при ~ 240 кДа, и даже при такой большой элементарной ячейке и уширении пиков перекрытие пятен не наблюдалось и не препятствовало определению структуры (14).Хотя расширение дифракционных пятен можно ограничить с помощью энергетического фильтра, это происходит за счет уменьшения передаваемого сигнала. Тем не менее, использование энергетического фильтра может улучшить соотношение сигнал / шум в экспериментах MicroED, и влияние толстых кристаллов с большими элементарными ячейками на качество дифракции требует дальнейшего изучения.
При определении белковых структур из кристаллов толщиной до 1 микрометра с использованием как фазового извлечения ab initio , так и молекулярного замещения, мы демонстрируем, что эксперименты MicroED в конечном итоге ограничены эффектами поглощения и качеством кристаллов.В настоящее время мы оцениваем кристаллы ~ 500 нм или меньше, чтобы обеспечить максимальное качество данных (~ 2x средняя длина свободного пробега для 200 кВ), но структуры из более толстых кристаллов не обязательно исключаются. Фактически, наши результаты для даже более толстых кристаллов обеспечивают уверенность в том, что наблюдаемая дифракция может быть свободна от множественных артефактов рассеяния в типичном эксперименте MicroED с непрерывным вращением, даже на кристаллах белка микрометровой толщины.
Заключительные замечания
Мы демонстрируем, что MicroED дает точные значения интенсивности дифракции, и что структуры могут быть определены с помощью молекулярного замещения или методов ab initio из кристаллов белка, намного более толстых, чем предполагалось ранее.Наши данные демонстрируют, что, как и в случае дифракции рентгеновских лучей, динамическое рассеяние не является недопустимым источником ошибок, когда данные MicroED собираются на кристаллах белка путем непрерывного вращения. Вместо этого явления поглощения ограничивают достижимое разрешение таким образом, что его можно обуздать путем объединения нескольких наборов данных. Мы предлагаем жесткую верхнюю границу толщины кристалла около одного микрометра в экспериментах с MicroED и мягкую верхнюю границу около 500 нм для обеспечения высококачественной дифракции с наилучшим возможным разрешением; эти ограничения налагаются не динамическим рассеянием, а поглощением.Наше исследование расширяет полезность MicroED как общего метода определения структуры до атомного разрешения от образцов до микрометра, открывая новые направления исследований, которые могут широко повлиять на структурную биологию.
Материалы и методы
Препарат белка
hPrP-β2α2 с чистотой более 98% был приобретен у Genscript, растворен в воде в концентрации 10-20 мг / мл и подвергнут скринингу методом «висящей капли» на высокопроизводительном сите. Первоначальные попадания были оптимизированы в 24-луночных висячих лотках для капель.Лучшие кристаллы наблюдались в условиях, содержащих 10% (мас. / Об.) ПЭГ-8000; 0,1 М MES pH 6,0; Zn (OAc) 2. Это условие было использовано в качестве основы для периодической кристаллизации пептида с концентрацией 10 мг / мл при соотношении раствора пептида к маточному раствору 1: 1. В этом состоянии кристаллы росли в виде кластеров игл, которые можно было сломать силой пипетирования и приложить к решеткам для криоконсервации. Протеиназа К ( E. Album ) была приобретена у Sigma и использовалась без дополнительной очистки. Кристаллы выращивали добавлением 5 мкл раствора белка (50 мг / мл) к 5 мкл раствора осадителя (1.5 M сульфат аммония, 0,1 M трис, pH 8,0) в лотке для диффузии паров сидячей капли. Большие кристаллы протеиназы собирали из сидящих капель и обрабатывали ультразвуком на более мелкие кристаллы, как описано ранее (4).
Сбор данных MicroED
Данные MicroED были собраны для 19 кристаллов пептида hPrP-β2α2 (Gly-Ser-Asn-Gln-Asn-Asn-Phe) различной толщины по реальному пространственному клину, соответствующему от -30 ° до +30 °. ° до при непрерывном вращении. Все эксперименты MicroED проводились на микроскопе FEI Tecnai F20 при ускоряющем напряжении 200 кВ, соответствующем длине волны 0.0251 Å. Данные были собраны с использованием постоянной скорости вращения 0,2 ° с -1 на CMOS-детекторе TVIPS TemCam-F416 в режиме рольставни с экспозицией 5 с. Интенсивность пучка для всех кристаллов hPrP-β2α2 была постоянной при средней мощности дозы 0,003e — Å -1 сек -1 или общей экспозиции ~ 1e — Å -2 . Кристаллы протеиназы К собирали при мощности дозы 0,01 e — Å -1 сек -1 с общей общей дозой на кристалл менее 3 e — Å -2 .Образцы hPrP-β2α2 поддерживали длину камеры 0,730 м, что эквивалентно расстоянию между образцом и детектором 1,313 м в соответствующей безлинзовой системе. Данные протеиназы К собирали с камеры длиной 1,2 м, что соответствует расстоянию детектора 2.200 м. Дифракционные данные собирали через круглую апертуру с выбранной площадью 1 мкм 2 в проекции для уменьшения фонового шума. Все измерения ПЭМ проводились при температурах жидкого азота (~ 77 К).
Определение толщины кристаллов
Толщина кристаллов оценивалась двумя способами: геометрическим путем и с помощью счета на камере.Во-первых, средняя толщина кристалла была первоначально оценена геометрически путем исследования площади проецирования каждого кристалла по изображениям, полученным при 0 °, 15 °, 30 °, 45 ° и 60 °, с использованием калибровки длины пикселя из известного размера отверстия сетки, равного 1 или 2 мкм в ImageJ (NIH). Кристаллы hPrP-β2α2 все имели форму стержней и предполагалось, что они являются стержнями эллипсоидальной формы. В серии наклона оценивалось соотношение сторон главной ( a ) и малой ( b ) оси под каждым углом, при известной длине 2 a ширина стержня, измеренная при 0 °.Средняя толщина кристалла составляет π / 4 = ~ 0,785% от максимальной толщины, 2 b , перпендикулярно пленке. Предполагалось, что кристаллы протеиназы К представляют собой кубоиды с длиной ребер w , l , h . Длины кромок w и l были найдены из изображений 0 °, а кромка h была оценена по изменению площади проекции кристалла на фиксированной длине вдоль оси x и y . Средняя толщина этих кристаллов принималась равной оценке края h .Точность измерения составляла от 5 до 10 нм, средняя дисперсия для каждого кристалла составляла ~ 20%, а стандартное отклонение между оценками интенсивности и геометрии составляло ~ 100 нм. Во-вторых, геометрическое приближение толщины кристалла, приведенное выше, было подтверждено с использованием соотношений интенсивностей, как описано ранее (2, 14, 26). Каждый кристалл из набора hPrP-β2α2 в Рисунок-3 / SI Document 2 при 0 ° были идентифицированы при 0 °. У каждого кристалла была область, выбранная из той части кристалла, на которой была рассчитана средняя передаваемая интенсивность.Затем ту же площадь использовали для расчета средней интенсивности для ближайшей области чистого углерода и ближайшего пустого отверстия сетки для сравнения. Толщина кристалла была оценена путем первой оценки толщины углерода с использованием формулы, приведенной в Feja & Aebi (26), в предположении, что вся недостающая интенсивность была потеряна из-за сигналов потери энергии, а максимальная интенсивность падающего излучения соответствовала интенсивности луча в отверстии решетки. Средняя длина свободного пробега аморфного углерода была рассчитана с использованием средней потери энергии углерода, равной 14.1 эВ, полученная по формуле, приведенной в (26), с Z = 6, что соответствует длине свободного пробега 125 нм при 200 кВ. Это дало постоянное значение толщины углерода примерно 45 нм со стандартным отклонением 7 нм. Толщина кристалла затем рассчитывается путем вычитания вклада углерода с использованием того же уравнения со средней длиной свободного пробега 242 нм, рассчитанной путем масштабирования значения 203 нм средней длины свободного пробега стекловидного льда, приведенного Grimm et al. на 120кВ (25, 30). Было обнаружено, что наши две меры имеют стандартное отклонение примерно 100 нм.Коэффициент корреляции между двумя показателями составил 89%. Это говорит о том, что наши геометрические измерения имеют точность в пределах 25% от перечисленных нами значений. График корреляции и отклонения от этих показателей интенсивности приведены на рис. 6 SI .
Эксперименты по поглощению
Энергетические спектры были получены на микроскопе JEOL JEM-3200FSC при 300 кВ с апертурой 500 нм 2 в проекции и дозой 0,1 э. — общая Å -2 .Спектры собирали на CMOS-детекторе TVIPS TemCam-F416 при полном разрешении в нормальном (интегрированном) режиме и сохраняли в виде 16-битных целочисленных файлов TIFF со знаком. Спектры были масштабированы в ImageJ до известного шага энергетического фильтра 100 эВ мкм -1 и размера пикселя 15,6 мкм. Отдельные изображения были загружены в систему Mathematica и выровнены по их главному компоненту по горизонтальной оси, что обычно приводило к повороту на 6,7 ° по часовой стрелке. Линейные сканы шириной 1 пиксель выбирались через пик с нулевыми потерями вдоль главной оси потерь и смещались таким образом, чтобы их пиковые интенсивности располагались при потерях энергии 0 эВ.Толщина кристаллов, используемых в спектрах, определялась, как описано выше. Пики с нулевыми потерями соответствовали общей гауссовой модели I = a * e -b (x-c) 2 для определения максимумов пиков и интегральной пиковой интенсивности. Точки данных были взвешены по квадратному корню из их интенсивностей для подгонки, чтобы гарантировать правильные решения. Интенсивность пика с нулевыми потерями соответствовала общей модели экспоненциального затухания с точкой пересечения, фиксированной на нормированном значении пика вакуума, как на , рис. 4, . Экспоненциальная аппроксимация экспериментальных данных для углеродной пленки, протеиназы K и кристаллов белка hPrP-β2α2 привела к остатку R 2 , равному 1.0, 0,85 и 0,88 соответственно.
Обработка данных MicroED
Дифракционные фильмы были преобразованы в формат файла SMV с помощью инструментов TVIPS (31) и проверены на усечение пикселей, как описано ранее (21). Индексирование и интеграция выполнялись в XDS (20, 32). Интегральные интенсивности дифракции были отсортированы и объединены в XSCALE. Объединенные интенсивности были преобразованы в амплитуды, а файлы отформатированы в формат SHELX в XDSCONV. Толстый и тонкий hPrP-β2α2 были решены с помощью SHELXT, правильно разместив все атомы в элементарной ячейке (33).SHELXT поддержал гипотезу кристаллографической симметрии P1 (16). Интенсивности протеиназы К вводились непосредственно из XSCALE в Phaser для молекулярного замещения (34). Молекулярная замена была успешной с LLG> 1000 и TFZ> 20 с использованием модели 5i9s . Уточнение проводилось с помощью phenix.refine (22). Индивидуальные модели были скорректированы вручную в Coot (35) путем визуального осмотра атомной модели по картам F o -F c и 2F o -F c .Всеатомные композитные карты пропусков с моделированием отжига были рассчитаны в Phenix (22).
Модели и рисунки
Рисунки были созданы либо в PyMol, либо в VMD (36, 37). Графики и подгонки к таким данным, как спектры EELS, были созданы в системе Mathematica с использованием нелинейных подгонок модели к данным. RMSD магистрали рассчитывались в VMD. Все значения RMSD атомов были сгенерированы командой align в PyMol.
Благодарности
Мы благодарим Дэвида Эйзенберга (HHMI, UCLA), Роберта Глезера (UCB) и Майкла Савайя (UCLA) за полезные обсуждения и / или критическое чтение этой рукописи.Хосе Родригеса поддерживают как стипендиата Серла и молодого исследователя Бекмана. Лаборатория Гонена поддерживается Медицинским институтом Говарда Хьюза. Эта работа также была поддержана программой посетителей-исследователей Janelia.
Контролируемое складывание структур микрометрового размера на JSTOR
АбстрактныйБыло изготовлено несколько типов микроактюаторов, от простых лопастей до самосборных и разборных кубиков. Проводящие бислои из слоя полимера и слоя золота использовались в качестве шарниров для соединения жестких пластин друг с другом и с кремниевой подложкой.Изгиб петель был электрически управляемым и обратимым, что позволяло точно позиционировать пластины в трехмерном пространстве. Структуры были освобождены от подложки с помощью техники, основанной на дифференциальной адгезии. Этот метод, который позволяет избежать использования защитного слоя и позволяет исполнительным механизмам отрываться от поверхности, может иметь общие применения в микрообработке. Возможности включают изготовление поверхностей, светоотражение или химические свойства которых можно переключать.
Информация о журналеScience, основанный Томасом А. Эдисоном в 1880 году и издаваемый AAAS, сегодня является крупнейшим в мире общенаучным журналом с тиражом. Публикуемый 51 раз в год, журнал Science известен своими высоко цитируемыми, рецензируемыми научными работами, своей особой силой в дисциплинах наук о жизни и отмеченным наградами освещением последних научных новостей. Онлайн-издание включает не только полный текст текущих выпусков, но и научные архивы, относящиеся к первому изданию Эдисона в 1880 году.Сайт Science Careers, который можно найти в печати и в Интернете, предоставляет еженедельно публикуемые статьи о карьере, тысячи объявлений о вакансиях, обновляемых несколько раз в неделю, и другие услуги, связанные с карьерой. В интерактивном научном мультимедийном центре представлены научные подкасты, изображения и слайд-шоу, видео, семинары и другие интерактивные функции. Для получения дополнительной информации посетите www.sciencemag.org.
Информация об издателеAAAS, основанная в 1848 году, превратилась в крупнейшее в мире междисциплинарное научное общество, насчитывающее почти 130 000 членов и подписчиков.Миссия «продвигать науку, технику и инновации во всем мире на благо всех людей» вывела организацию на передний план национальных и международных инициатив. Глобальные усилия включают программы и партнерства по всему миру, от Азии до Европы и Африки, а также обширную работу в области прав человека с использованием геопространственных технологий для подтверждения нарушений. Программы по науке и политике включают в себя крупный ежегодный форум по политике в области науки и технологий, стипендии в рамках политики в области науки и технологий в Конгрессе США и правительственных агентствах, а также отслеживание финансирования США исследований в области НИОКР.Инициативы в области естественнонаучного образования заложили основу для обучения на основе стандартов и предоставляют учителям инструменты поддержки в Интернете.